

The extended transmembrane Orai1 N-terminal (ETON) region combines binding interface and gate for Orai1 activation by STIM1
- PMID: 23943619
- PMCID: PMC3790000
- DOI: 10.1074/jbc.M113.501510
The extended transmembrane Orai1 N-terminal (ETON) region combines binding interface and gate for Orai1 activation by STIM1
Abstract
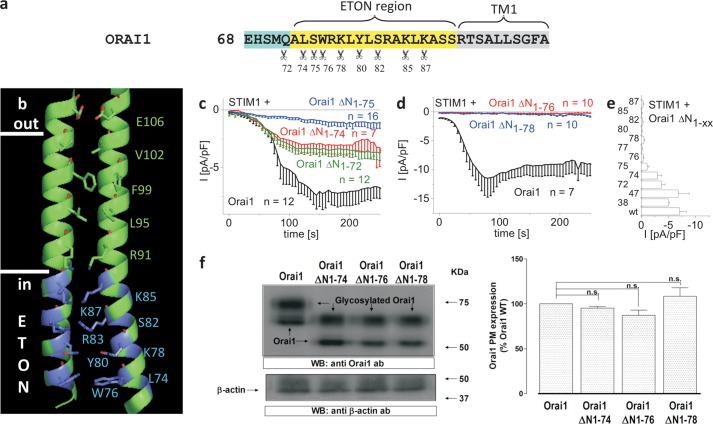
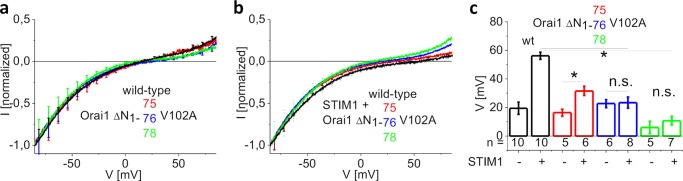
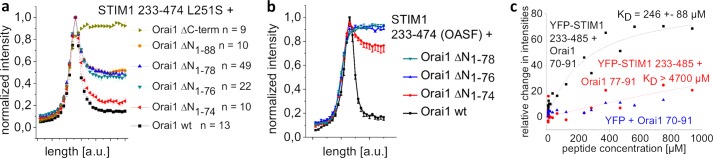
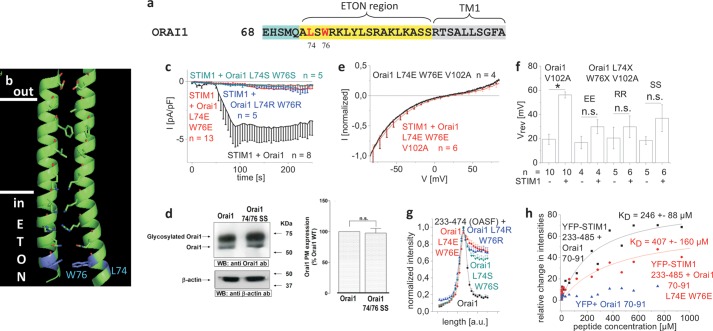
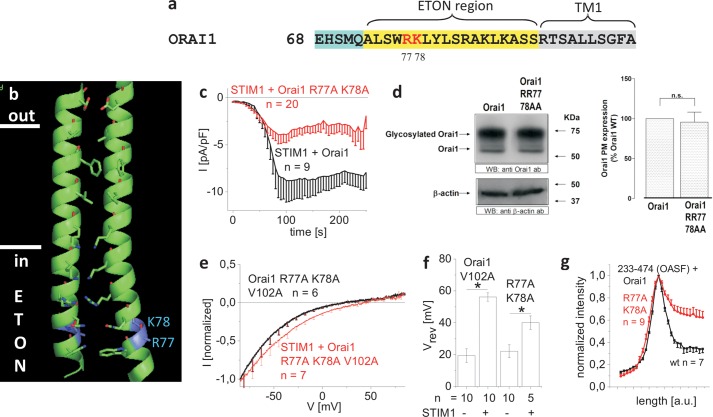
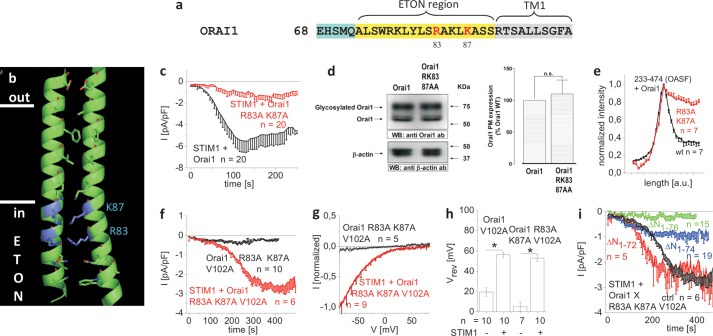
STIM1 and Orai1 represent the two molecular key components of the Ca(2+) release-activated Ca(2+) channels. Their activation involves STIM1 C terminus coupling to both the N terminus and the C terminus of Orai. Here we focused on the extended transmembrane Orai1 N-terminal (ETON, aa73-90) region, conserved among the Orai family forming an elongated helix of TM1 as recently shown by x-ray crystallography. To identify "hot spot" residues in the ETON binding interface for STIM1 interaction, numerous Orai1 constructs with N-terminal truncations or point mutations within the ETON region were generated. N-terminal truncations of the first four residues of the ETON region or beyond completely abolished STIM1-dependent Orai1 function. Loss of Orai1 function resulted from neither an impairment of plasma membrane targeting nor pore damage, but from a disruption of STIM1 interaction. In a complementary approach, we monitored STIM1-Orai interaction via Orai1 V102A by determining restored Ca(2+) selectivity as a consequence of STIM1 coupling. Orai1 N-terminal truncations that led to a loss of function consistently failed to restore Ca(2+) selectivity of Orai1 V102A in the presence of STIM1, demonstrating impairment of STIM1 binding. Hence, the major portion of the ETON region (aa76-90) is essential for STIM1 binding and Orai1 activation. Mutagenesis within the ETON region revealed several hydrophobic and basic hot spot residues that appear to control STIM1 coupling to Orai1 in a concerted manner. Moreover, we identified two basic residues, which protrude into the elongated pore to redound to Orai1 gating. We suggest that several hot spot residues in the ETON region contribute in aggregate to the binding of STIM1, which in turn is coupled to a conformational reorientation of the gate.
Keywords: Calcium Channels; Calcium Intracellular Release; Calcium Signaling; Electrophysiology; Ion Channels.
Figures
Similar articles
-
Differential roles of the C and N termini of Orai1 protein in interacting with stromal interaction molecule 1 (STIM1) for Ca2+ release-activated Ca2+ (CRAC) channel activation.J Biol Chem. 2013 Apr 19;288(16):11263-72. doi: 10.1074/jbc.M113.450254. Epub 2013 Feb 27. J Biol Chem. 2013. PMID: 23447534 Free PMC article.
-
Cooperative Binding of Stromal Interaction Molecule 1 (STIM1) to the N and C Termini of Calcium Release-activated Calcium Modulator 1 (Orai1).J Biol Chem. 2016 Jan 1;291(1):334-41. doi: 10.1074/jbc.M115.685289. Epub 2015 Nov 6. J Biol Chem. 2016. PMID: 26546674 Free PMC article.
-
SOAR and the polybasic STIM1 domains gate and regulate Orai channels.Nat Cell Biol. 2009 Mar;11(3):337-43. doi: 10.1038/ncb1842. Epub 2009 Feb 1. Nat Cell Biol. 2009. PMID: 19182790 Free PMC article.
-
Store-Independent Orai Channels Regulated by STIM.In: Kozak JA, Putney JW Jr, editors. Calcium Entry Channels in Non-Excitable Cells. Boca Raton (FL): CRC Press/Taylor & Francis; 2018. Chapter 11. In: Kozak JA, Putney JW Jr, editors. Calcium Entry Channels in Non-Excitable Cells. Boca Raton (FL): CRC Press/Taylor & Francis; 2018. Chapter 11. PMID: 30299650 Free Books & Documents. Review.
-
The STIM-Orai Pathway: Conformational Coupling Between STIM and Orai in the Activation of Store-Operated Ca2+ Entry.Adv Exp Med Biol. 2017;993:83-98. doi: 10.1007/978-3-319-57732-6_5. Adv Exp Med Biol. 2017. PMID: 28900910 Free PMC article. Review.
Cited by
-
ER-resident STIM1/2 couples Ca2+ entry by NMDA receptors to pannexin-1 activation.Proc Natl Acad Sci U S A. 2022 Sep 6;119(36):e2112870119. doi: 10.1073/pnas.2112870119. Epub 2022 Aug 29. Proc Natl Acad Sci U S A. 2022. PMID: 36037373 Free PMC article.
-
Store-operated calcium entry: Mechanisms and modulation.Biochem Biophys Res Commun. 2015 Apr 24;460(1):40-9. doi: 10.1016/j.bbrc.2015.02.110. Biochem Biophys Res Commun. 2015. PMID: 25998732 Free PMC article. Review.
-
Store-Operated Calcium Entry in the Cardiovascular System.Adv Exp Med Biol. 2021;1349:303-333. doi: 10.1007/978-981-16-4254-8_14. Adv Exp Med Biol. 2021. PMID: 35138620 Review.
-
Orai1 Boosts SK3 Channel Activation.Cancers (Basel). 2021 Dec 17;13(24):6357. doi: 10.3390/cancers13246357. Cancers (Basel). 2021. PMID: 34944977 Free PMC article.
-
STIM1 triggers a gating rearrangement at the extracellular mouth of the ORAI1 channel.Nat Commun. 2014 Oct 9;5:5164. doi: 10.1038/ncomms6164. Nat Commun. 2014. PMID: 25296861 Free PMC article.
References
-
- Prakriya M., Feske S., Gwack Y., Srikanth S., Rao A., Hogan P. G. (2006) Orai1 is an essential pore subunit of the CRAC channel. Nature 443, 230–233 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
