

Phospholipase B is activated in response to sterol removal and stimulates acrosome exocytosis in murine sperm
- PMID: 23943622
- PMCID: PMC3784722
- DOI: 10.1074/jbc.M113.450981
Phospholipase B is activated in response to sterol removal and stimulates acrosome exocytosis in murine sperm
Abstract
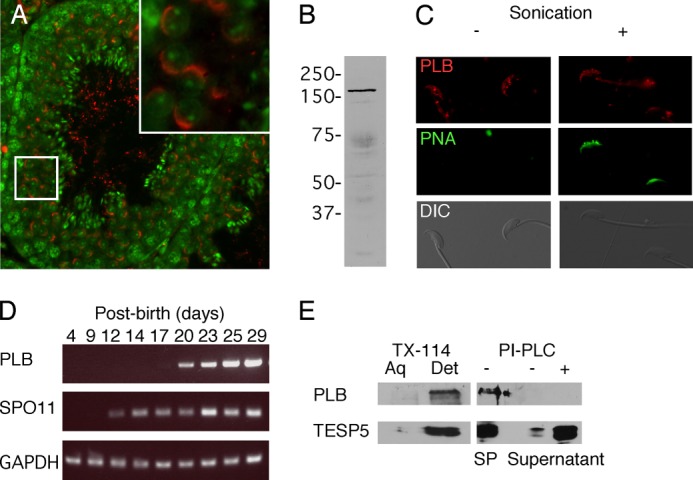
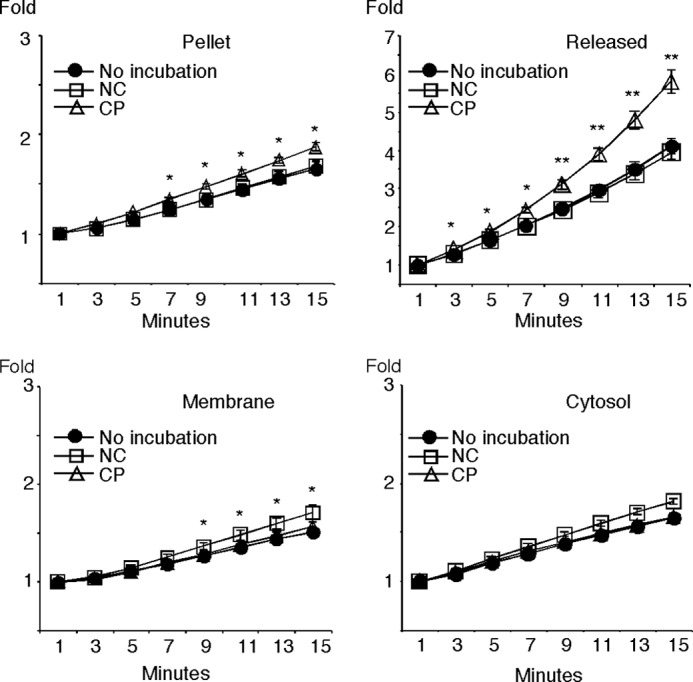
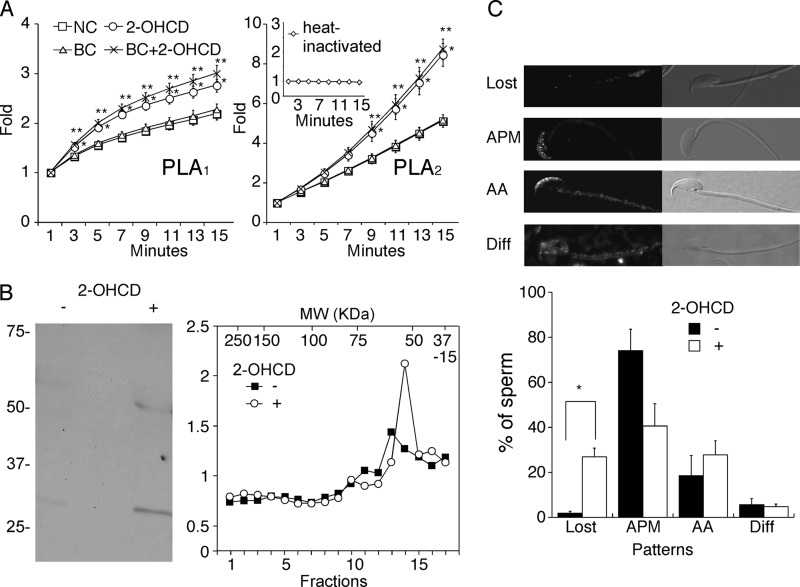
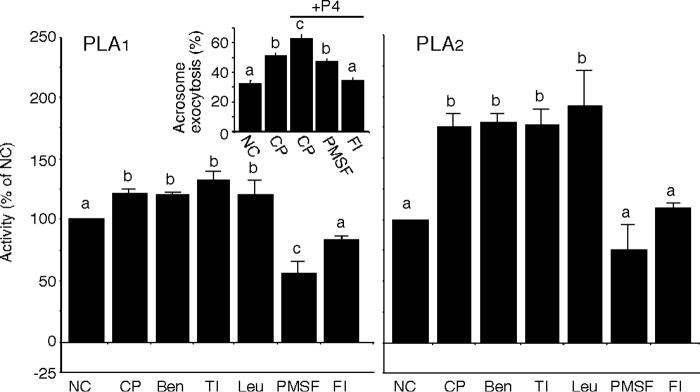
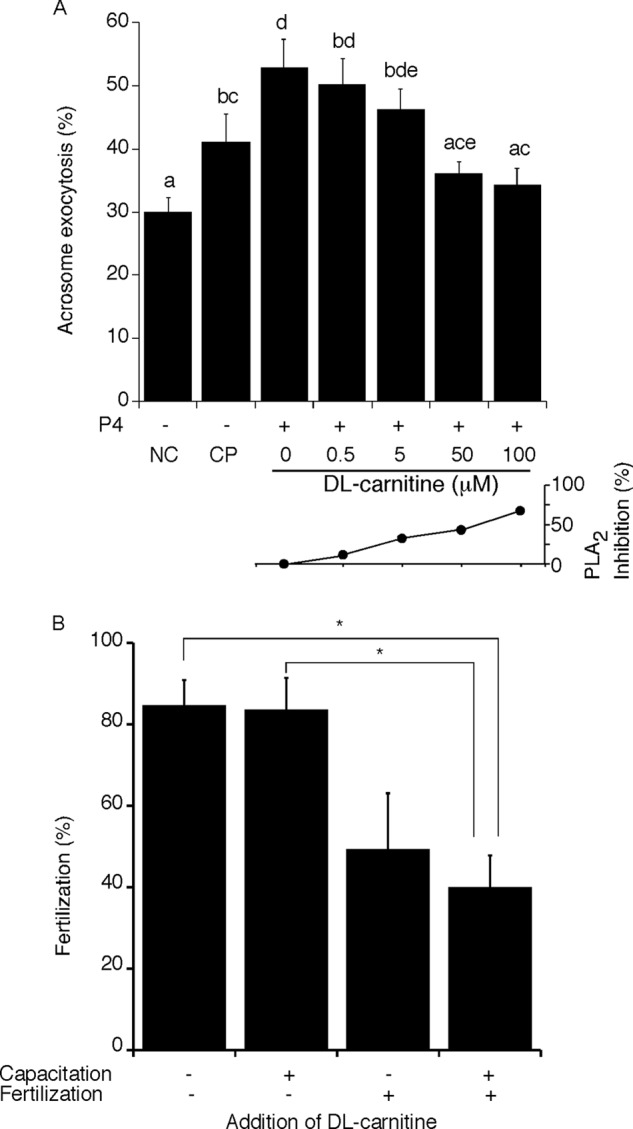
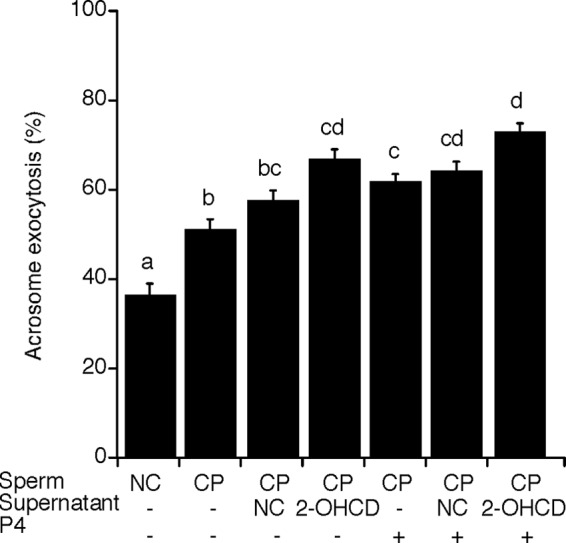
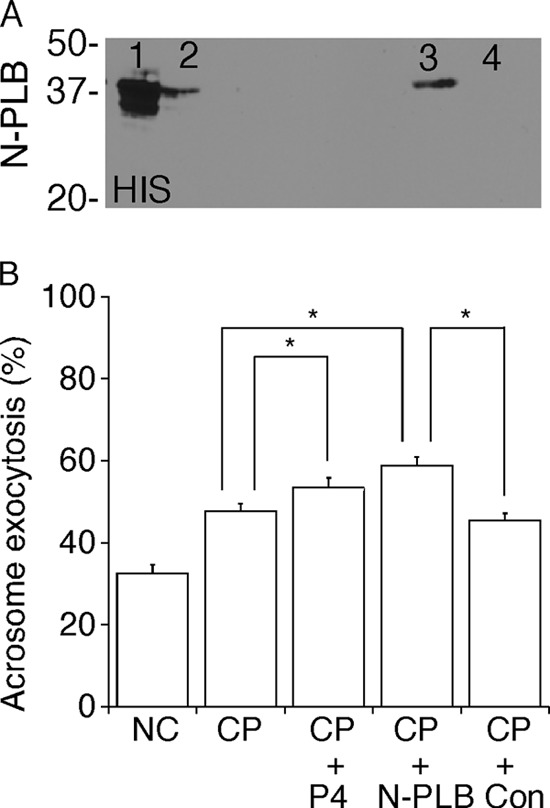
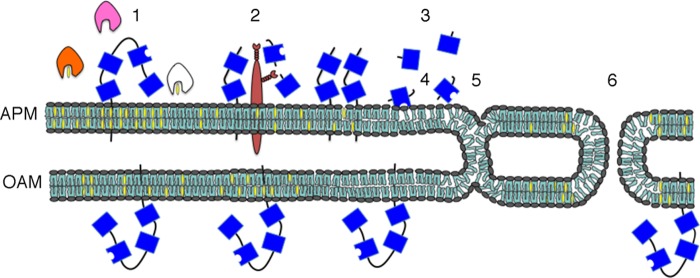
Despite a strict requirement for sterol removal for sperm to undergo acrosome exocytosis (AE), the mechanisms by which changes in membrane sterols are transduced into changes in sperm fertilization competence are poorly understood. We have previously shown in live murine sperm that the plasma membrane overlying the acrosome (APM) contains several types of microdomains known as membrane rafts. When characterizing the membrane raft-associated proteomes, we identified phospholipase B (PLB), a calcium-independent enzyme exhibiting multiple activities. Here, we show that sperm surface PLB is activated in response to sterol removal. Both biochemical activity assays and immunoblots of subcellular fractions of sperm incubated with the sterol acceptor 2-hydroxypropyl-β-cyclodextrin (2-OHCD) confirmed the release of an active PLB fragment. Specific protease inhibitors prevented PLB activation, revealing a mechanistic requirement for proteolytic cleavage. Competitive inhibitors of PLB reduced the ability of sperm both to undergo AE and to fertilize oocytes in vitro, suggesting an important role in fertilization. This was reinforced by our finding that incubation either with protein concentrate released from 2-OHCD-treated sperm or with recombinant PLB peptide corresponding to the catalytic domain was able to induce AE in the absence of other stimuli. Together, these results lead us to propose a novel mechanism by which sterol removal promotes membrane fusogenicity and AE, helping confer fertilization competence. Importantly, this mechanism provides a basis for the newly emerging model of AE in which membrane fusions occur during capacitation/transit through the cumulus, prior to any physical contact between the sperm and the oocyte's zona pellucida.
Keywords: Acrosome Exocytosis; Acrosome Reaction; Capacitation; Fertilization; Membrane Lipids; Phospholipase B; Raft; Serine Protease; Sperm.
Figures
Similar articles
-
Membrane rafts regulate phospholipase B activation in murine sperm.Commun Integr Biol. 2013 Nov 1;6(6):e27362. doi: 10.4161/cib.27362. Epub 2013 Dec 17. Commun Integr Biol. 2013. PMID: 24753791 Free PMC article.
-
Lipid Regulation of Acrosome Exocytosis.Adv Anat Embryol Cell Biol. 2016;220:107-27. doi: 10.1007/978-3-319-30567-7_6. Adv Anat Embryol Cell Biol. 2016. PMID: 27194352 Review.
-
Mouse sperm begin to undergo acrosomal exocytosis in the upper isthmus of the oviduct.Dev Biol. 2016 Mar 15;411(2):172-182. doi: 10.1016/j.ydbio.2016.02.006. Epub 2016 Feb 10. Dev Biol. 2016. PMID: 26872876 Free PMC article.
-
Most fertilizing mouse spermatozoa begin their acrosome reaction before contact with the zona pellucida during in vitro fertilization.Proc Natl Acad Sci U S A. 2011 Mar 22;108(12):4892-6. doi: 10.1073/pnas.1018202108. Epub 2011 Mar 7. Proc Natl Acad Sci U S A. 2011. PMID: 21383182 Free PMC article.
-
Capacitation-dependent reorganization of microdomains in the apical sperm head plasma membrane: functional relationship with zona binding and the zona-induced acrosome reaction.Theriogenology. 2008 Nov;70(8):1188-96. doi: 10.1016/j.theriogenology.2008.06.021. Epub 2008 Jul 21. Theriogenology. 2008. PMID: 18640708 Review.
Cited by
-
Comparison of Membrane Characteristics between Freshly Ejaculated and Cryopreserved Sperm in the Chicken.J Poult Sci. 2016 Oct 25;53(4):305-312. doi: 10.2141/jpsa.0160043. J Poult Sci. 2016. PMID: 32908398 Free PMC article.
-
Validation of a laboratory-developed test of human sperm capacitation.Mol Reprod Dev. 2017 May;84(5):408-422. doi: 10.1002/mrd.22801. Epub 2017 Apr 18. Mol Reprod Dev. 2017. PMID: 28418600 Free PMC article.
-
A genetically targeted sensor reveals spatial and temporal dynamics of acrosomal calcium and sperm acrosome exocytosis.J Biol Chem. 2022 May;298(5):101868. doi: 10.1016/j.jbc.2022.101868. Epub 2022 Mar 27. J Biol Chem. 2022. PMID: 35346690 Free PMC article.
-
Theoretical Analysis of S, M and N Structural Proteins by the Protein-RNA Recognition Code Leads to Genes/proteins that Are Relevant to the SARS-CoV-2 Life Cycle and Pathogenesis.Front Genet. 2021 Sep 29;12:763995. doi: 10.3389/fgene.2021.763995. eCollection 2021. Front Genet. 2021. PMID: 34659373 Free PMC article.
-
Ultrastructure and lipid composition of detergent-resistant membranes derived from mammalian sperm and two types of epithelial cells.Cell Tissue Res. 2016 Jan;363(1):129-145. doi: 10.1007/s00441-015-2272-y. Epub 2015 Sep 16. Cell Tissue Res. 2016. PMID: 26378009 Free PMC article.
References
-
- Chang M. C. (1951) Fertilizing Capacity of spermatozoa deposited into the fallopian tubes. Nature 168, 697–698 - PubMed
-
- Austin C. R. (1951) Observations on the penetration of the sperm in the mammalian egg. Aust. J. Sci. Res. B 4, 581–596 - PubMed
-
- Visconti P. E., Bailey J. L., Moore G. D., Pan D., Olds-Clarke P., Kopf G. S. (1995) Capacitation of mouse spermatozoa. I. Correlation between the capacitation state and protein tyrosine phosphorylation. Development 121, 1129–1137 - PubMed
-
- Kim K. S., Gerton G. L. (2003) Differential release of soluble and matrix components. Evidence for intermediate states of secretion during spontaneous acrosomal exocytosis in mouse sperm. Dev. Biol. 264, 141–152 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
