

Adipocyte fetuin-A contributes to macrophage migration into adipose tissue and polarization of macrophages
- PMID: 23943623
- PMCID: PMC3784748
- DOI: 10.1074/jbc.C113.495473
Adipocyte fetuin-A contributes to macrophage migration into adipose tissue and polarization of macrophages
Abstract
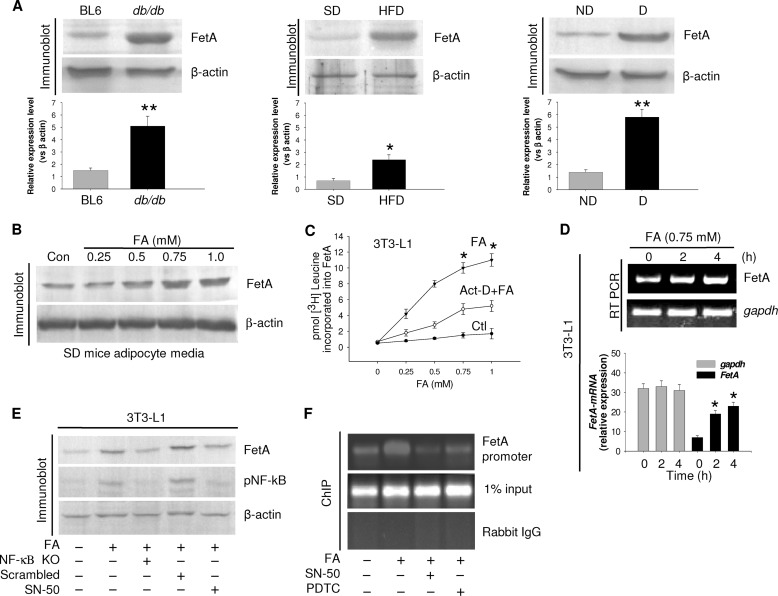
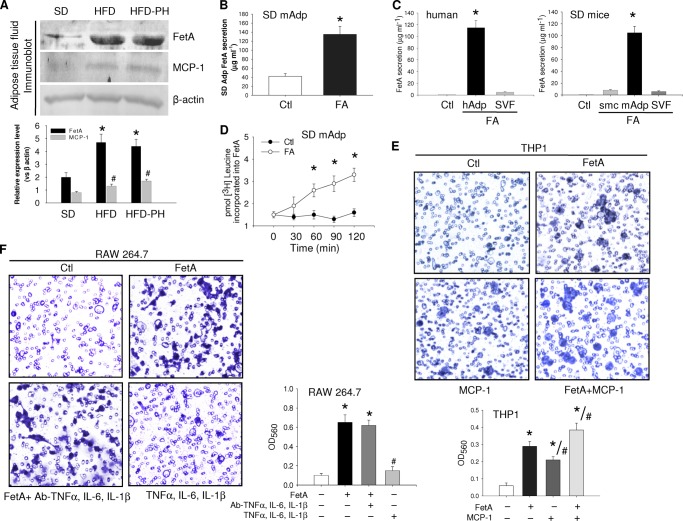
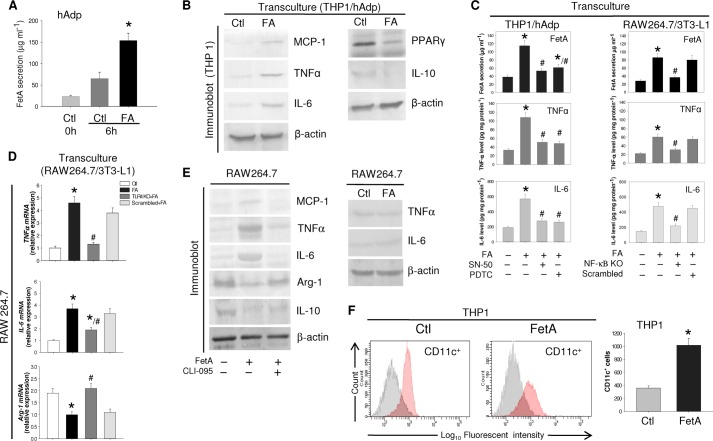
Macrophage infiltration into adipose tissue during obesity and their phenotypic conversion from anti-inflammatory M2 to proinflammatory M1 subtype significantly contributes to develop a link between inflammation and insulin resistance; signaling molecule(s) for these events, however, remains poorly understood. We demonstrate here that excess lipid in the adipose tissue environment may trigger one such signal. Adipose tissue from obese diabetic db/db mice, high fat diet-fed mice, and obese diabetic patients showed significantly elevated fetuin-A (FetA) levels in respect to their controls; partially hepatectomized high fat diet mice did not show noticeable alteration, indicating adipose tissue to be the source of this alteration. In adipocytes, fatty acid induces FetA gene and protein expressions, resulting in its copious release. We found that FetA could act as a chemoattractant for macrophages. To simulate lipid-induced inflammatory conditions when proinflammatory adipose tissue and macrophages create a niche of an altered microenvironment, we set up a transculture system of macrophages and adipocytes; the addition of fatty acid to adipocytes released FetA into the medium, which polarized M2 macrophages to M1. This was further confirmed by direct FetA addition to macrophages. Taken together, lipid-induced FetA from adipocytes is an efficient chemokine for macrophage migration and polarization. These findings open a new dimension for understanding obesity-induced inflammation.
Keywords: Adipose Tissue; Inflammation; Lipids; Macrophages; Obesity.
Figures
References
-
- Kahn S. E., Hull R. L., Utzschneider K. M. (2006) Mechanisms linking obesity to insulin resistance and type 2 diabetes. Nature 444, 840–846 - PubMed
-
- Gregor M. F., Hotamisligil G. S. (2011) Inflammatory mechanisms in obesity. Annu. Rev. Immunol. 29, 415–445 - PubMed
-
- Johnson A. M. F., Olefsky J. M. (2013) The origins and drivers of insulin resistance. Cell 152, 673–684 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
