

Old Cryptococcus neoformans cells contribute to virulence in chronic cryptococcosis
- PMID: 23943761
- PMCID: PMC3747583
- DOI: 10.1128/mBio.00455-13
Old Cryptococcus neoformans cells contribute to virulence in chronic cryptococcosis
Abstract
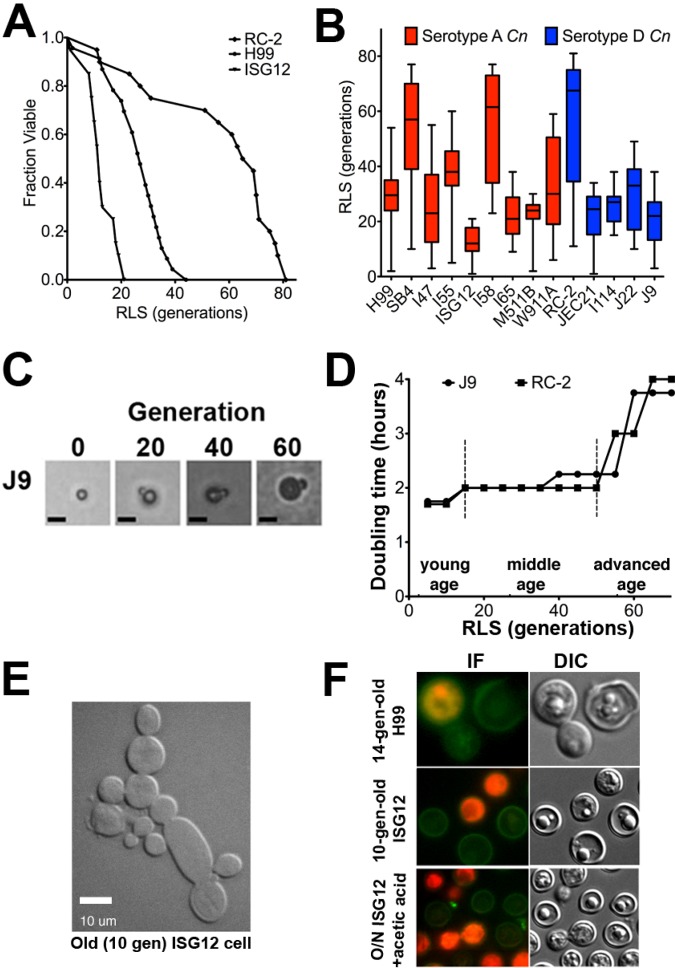
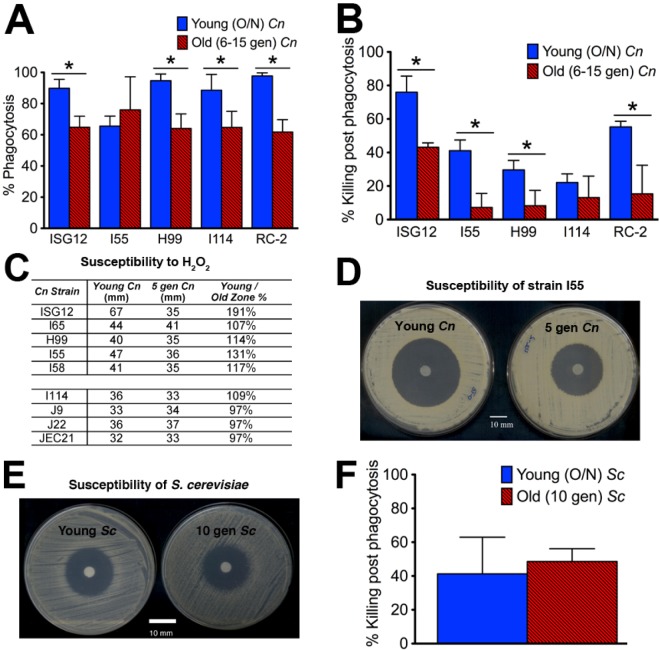
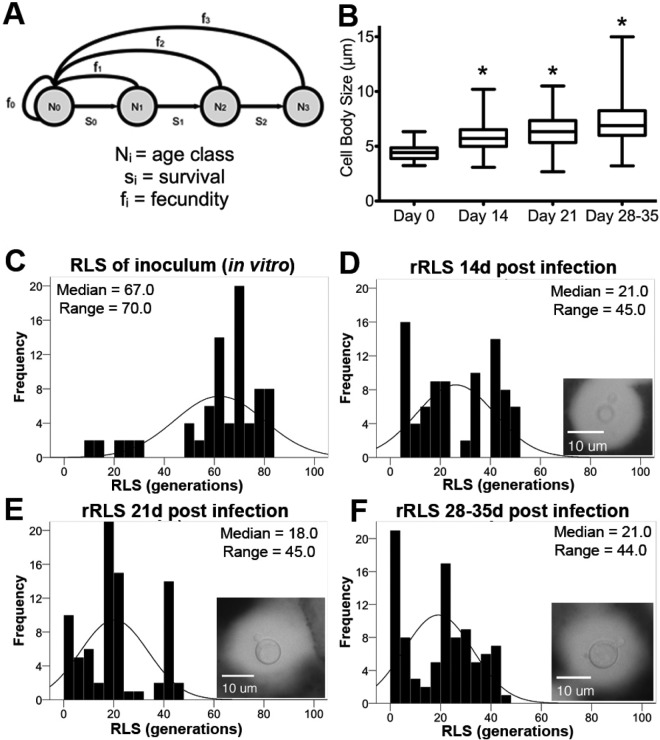
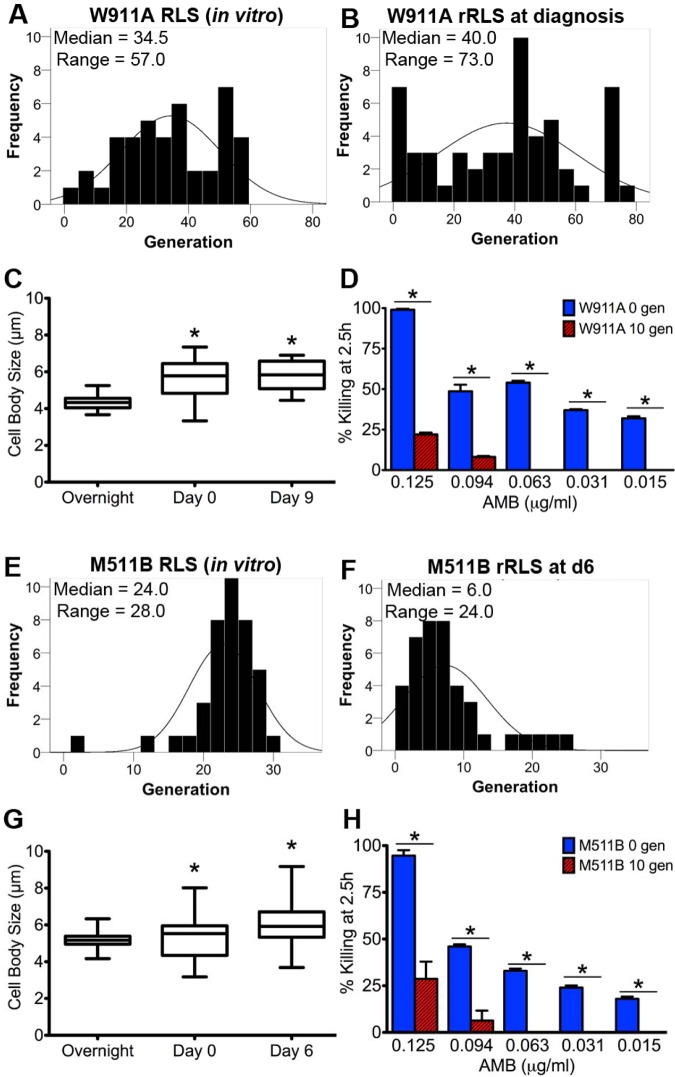
Does cell age matter in virulence? The emergence of persister cells during chronic infections is critical for persistence of infection, but little is known how this occurs. Here, we demonstrate for the first time that the replicative age of the fungal pathogen Cryptococcus neoformans contributes to persistence during chronic meningoencephalitis. Generationally older C. neoformans cells are more resistant to hydrogen peroxide stress, macrophage intracellular killing, and antifungal agents. Older cells accumulate in both experimental rat infection and in human cryptococcosis. Mathematical modeling supports the concept that the presence of older C. neoformans cells emerges from in vivo selection pressures. We propose that advanced replicative aging is a new unanticipated virulence trait that emerges during chronic fungal infection and facilitates persistence. Therapeutic interventions that target old cells could help in the clearance of chronic infections.
Importance: Our findings that the generational age of Cryptococcus neoformans cells matters in pathogenesis introduces a novel concept to eukaryotic pathogenesis research. We propose that emerging properties of aging C. neoformans cells and possibly also other fungal pathogens contribute to persistence and virulence. Whereas the replicative life span of strains may not matter for virulence per se, age-related resilience and thus the generational age of individual C. neoformans cells within a pathogen population could greatly affect persistence of the pathogen population and therefore impact outcome.
Figures
References
-
- Park BJ, Wannemuehler KA, Marston BJ, Govender N, Pappas PG, Chiller TM. 2009. Estimation of the current global burden of cryptococcal meningitis among persons living with HIV/AIDS. AIDS 23:525–530 - PubMed
-
- Manosuthi W, Sungkanuparph S, Thongyen S, Chumpathat N, Eampokalap B, Thawornwan U, Foongladda S. 2006. Antifungal susceptibilities of Cryptococcus neoformans cerebrospinal fluid isolates and clinical outcomes of cryptococcal meningitis in HIV-infected patients with/without fluconazole prophylaxis. J. Med. Assoc. Thai. 89:795–802 - PubMed
-
- Spitzer ED, Spitzer SG, Freundlich LF, Casadevall A. 1993. Persistence of initial infection in recurrent Cryptococcus neoformans meningitis. Lancet 341:595–596 - PubMed
-
- Bicanic T, Muzoora C, Brouwer AE, Meintjes G, Longley N, Taseera K, Rebe K, Loyse A, Jarvis J, Bekker LG, Wood R, Limmathurotsakul D, Chierakul W, Stepniewska K, White NJ, Jaffar S, Harrison TS. 2009. Independent association between rate of clearance of infection and clinical outcome of HIV-associated cryptococcal meningitis: analysis of a combined cohort of 262 patients. Clin. Infect. Dis. 49:702–709 - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
