

Acacetin inhibits in vitro and in vivo angiogenesis and downregulates Stat signaling and VEGF expression
- PMID: 23943785
- PMCID: PMC3808880
- DOI: 10.1158/1940-6207.CAPR-13-0209
Acacetin inhibits in vitro and in vivo angiogenesis and downregulates Stat signaling and VEGF expression
Abstract
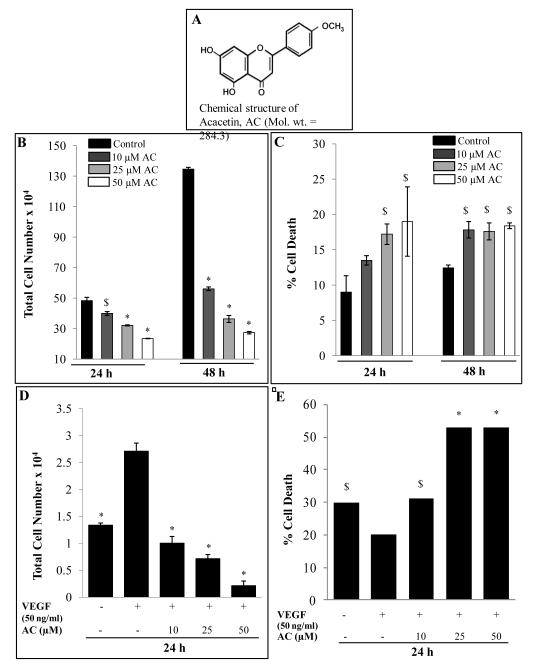
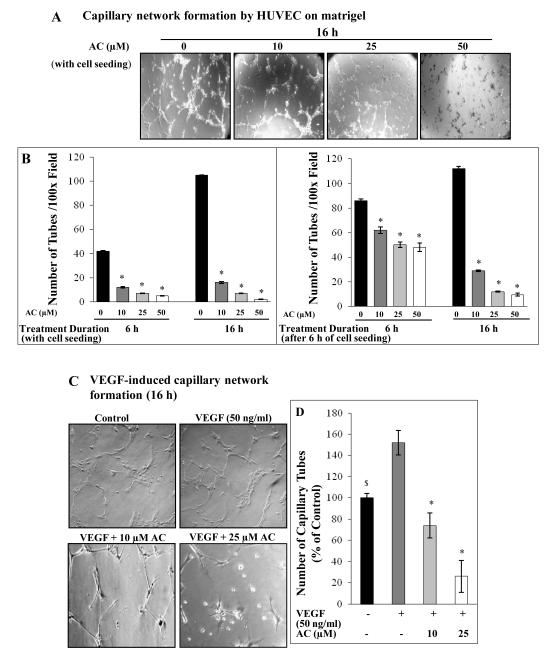
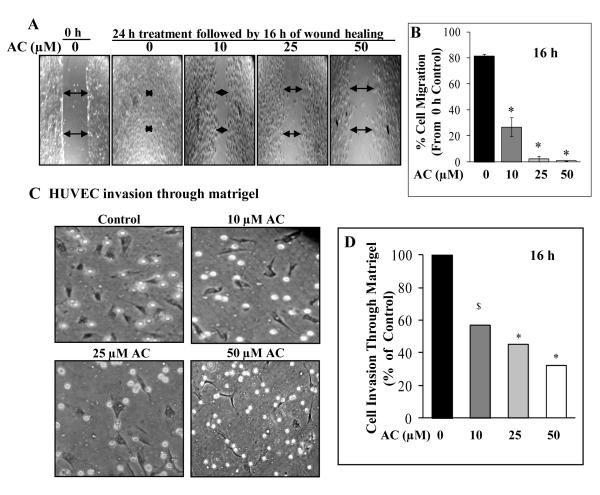
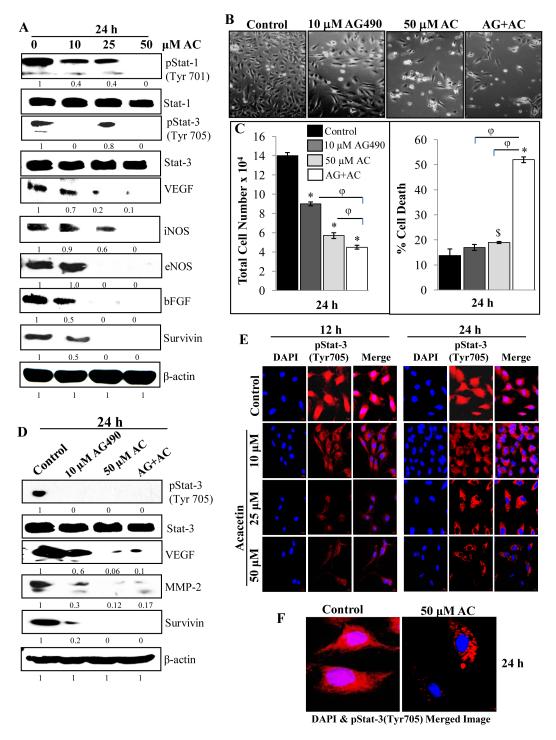
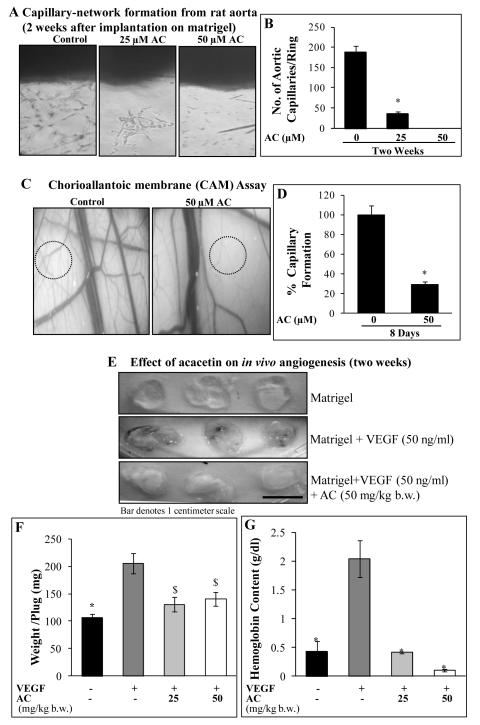
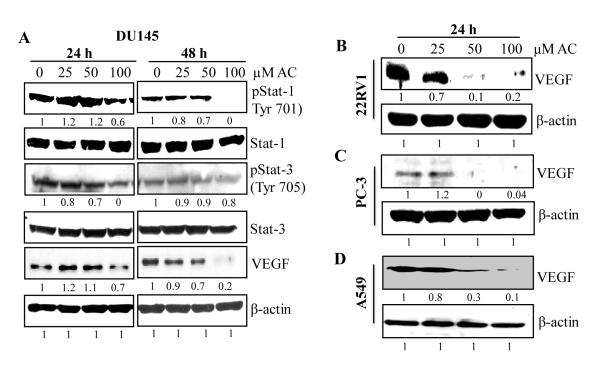
Angiogenesis is an effective target in cancer control. The antiangiogenic efficacy and associated mechanisms of acacetin, a plant flavone, are poorly known. In the present study, acacetin inhibited growth and survival (up to 92%; P < 0.001), and capillary-like tube formation on Matrigel (up to 98%; P < 0.001) by human umbilical vein endothelial cells (HUVEC) in regular condition, as well as VEGF-induced and tumor cells conditioned medium-stimulated growth conditions. It caused retraction and disintegration of preformed capillary networks (up to 91%; P < 0.001). HUVEC migration and invasion were suppressed by 68% to 100% (P < 0.001). Acacetin inhibited Stat-1 (Tyr701) and Stat-3 (Tyr705) phosphorylation, and downregulated proangiogenic factors including VEGF, endothelial nitric oxide synthase (eNOS), inducible nitric oxide synthase (iNOS), matrix metalloproteinase-2 (MMP-2), and basic fibroblast growth factor (bFGF) in HUVEC. It also suppressed nuclear localization of pStat-3 (Tyr705). Acacetin strongly inhibited capillary sprouting and networking from rat aortic rings and fertilized chicken egg chorioallantoic membrane (CAM; ∼71%; P < 0.001). Furthermore, it suppressed angiogenesis in Matrigel plugs implanted in Swiss albino mice. Acacetin also inhibited tyrosine phosphorylation of Stat-1 and -3, and expression of VEGF in cancer cells. Overall, acacetin inhibits Stat signaling and suppresses angiogenesis in vitro, ex vivo, and in vivo, and therefore, it could be a potential agent to inhibit tumor angiogenesis and growth.
Figures
References
-
- Ferrara N, Kerbel RS. Angiogenesis as a therapeutic target. Nature. 2005;438:967–74. - PubMed
-
- Naumov GN, Akslen LA, Folkman J. Role of angiogenesis in human tumor dormancy: animal models of the angiogenic switch. Cell Cycle. 2006;5(16):1779–87. - PubMed
-
- Naumov GN, Folkman J, Straume O. Tumor dormancy due to failure of angiogenesis: role of the microenvironment. Clin Exp Metastasis. 2009;26(1):51–60. - PubMed
-
- Halvorsen O, Haukaas S, Hoisaeter P, Akslen L. Independent prognostic importance of microvesel density in clinically localized prostate cancer. Anticancer Res. 2000;20:3791–99. - PubMed
-
- Gettman MT, Pacelli A, Slezak J, Bergstralh EJ, Blute M, Zincke H, Bostwick DG. Role of microvessel density in predicting recurrence in pathologic Stage T3 prostatic adenocarcinoma. Urology. 1999;54:479–85. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
