

Epithelial-mesenchymal transition and tumor suppression are controlled by a reciprocal feedback loop between ZEB1 and Grainyhead-like-2
- PMID: 23943797
- PMCID: PMC3806457
- DOI: 10.1158/0008-5472.CAN-12-4082
Epithelial-mesenchymal transition and tumor suppression are controlled by a reciprocal feedback loop between ZEB1 and Grainyhead-like-2
Abstract
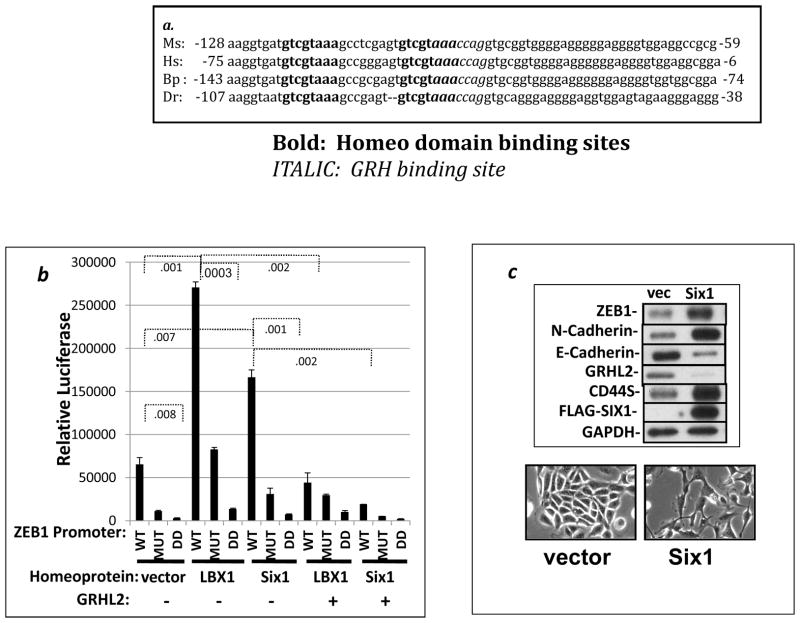
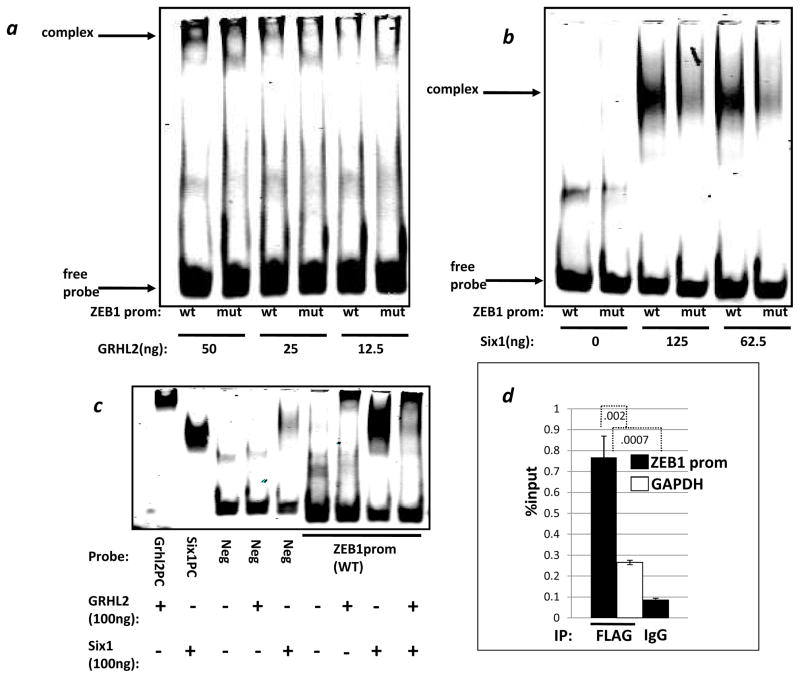
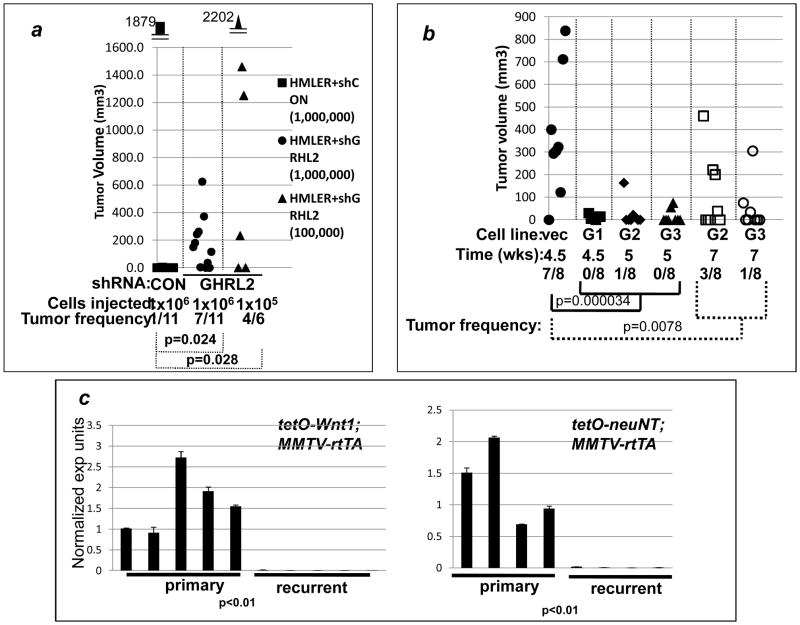
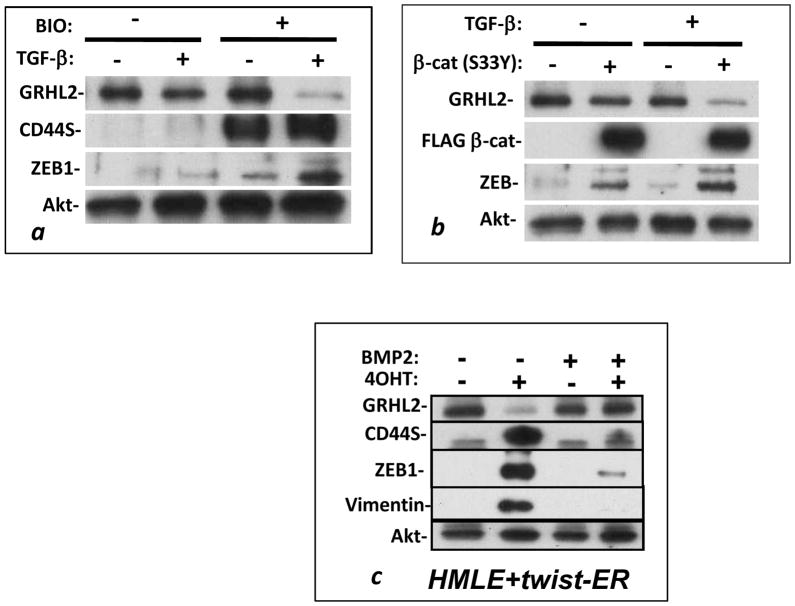
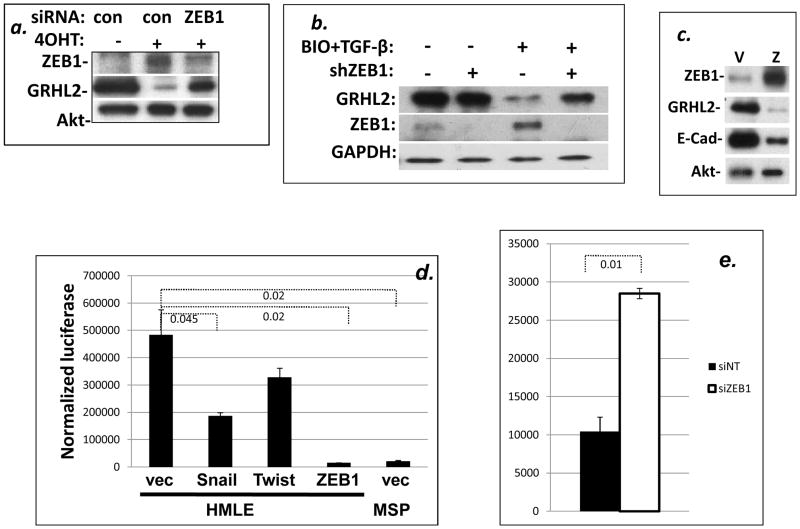
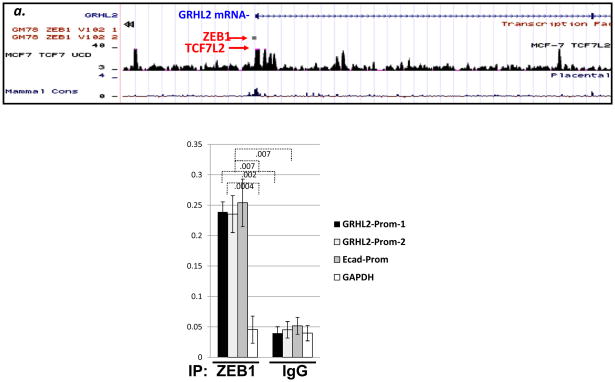
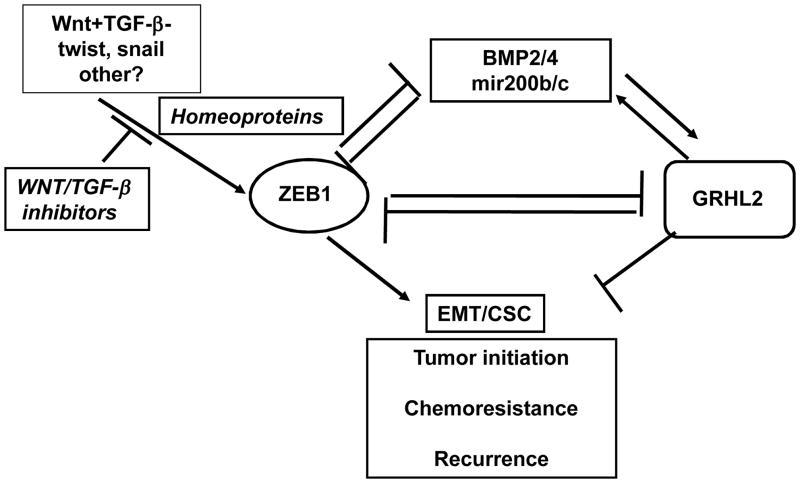
Epithelial-mesenchymal transition (EMT) in carcinoma cells enhances malignant progression by promoting invasion and survival. EMT is induced by microenvironmental factors, including TGF-β and Wnt agonists, and by the E-box-binding transcription factors Twist, Snail, and ZEB. Grainyhead-like-2 (GRHL2), a member of the mammalian Grainyhead family of wound-healing regulatory transcription factors, suppresses EMT and restores sensitivity to anoikis by repressing ZEB1 expression and inhibiting TGF-β signaling. In this study, we elucidate the functional relationship between GRHL2 and ZEB1 in EMT/MET and tumor biology. At least three homeodomain proteins, Six1, LBX1, and HoxA5, transactivated the ZEB1 promoter, in the case of Six1, through direct protein-promoter interaction. GRHL2 altered the Six1-DNA complex, inhibiting this transactivation. Correspondingly, GRHL2 expression prevented tumor initiation in xenograft assays, sensitized breast cancer cells to paclitaxel, and suppressed the emergence of CD44(high)CD24(low) cells (defining the cancer stem cell phenotype in the cell type studied). GRHL2 was downregulated in recurrent mouse tumors that had evolved to an oncogene-independent, EMT-like state, supporting a role for GRHL2 downregulation in this phenotypic transition, modeling disease recurrence. The combination of TGF-β and Wnt activation repressed GRHL2 expression by direct interaction of ZEB1 with the GRHL2 promoter, inducing EMT. Together, our observations indicate that a reciprocal feedback loop between GRHL2 and ZEB1 controls epithelial versus mesenchymal phenotypes and EMT-driven tumor progression.
©2013 AACR.
Conflict of interest statement
Figures
Similar articles
-
Suppression of the epithelial-mesenchymal transition by Grainyhead-like-2.Cancer Res. 2012 May 1;72(9):2440-53. doi: 10.1158/0008-5472.CAN-11-4038. Epub 2012 Feb 29. Cancer Res. 2012. PMID: 22379025 Free PMC article.
-
Dual roles of the transcription factor grainyhead-like 2 (GRHL2) in breast cancer.J Biol Chem. 2013 Aug 9;288(32):22993-3008. doi: 10.1074/jbc.M113.456293. Epub 2013 Jun 29. J Biol Chem. 2013. PMID: 23814079 Free PMC article.
-
The GRHL2/ZEB Feedback Loop-A Key Axis in the Regulation of EMT in Breast Cancer.J Cell Biochem. 2017 Sep;118(9):2559-2570. doi: 10.1002/jcb.25974. Epub 2017 May 3. J Cell Biochem. 2017. PMID: 28266048
-
Roles of Grainyhead-like transcription factors in cancer.Oncogene. 2017 Nov 2;36(44):6067-6073. doi: 10.1038/onc.2017.178. Epub 2017 Jul 17. Oncogene. 2017. PMID: 28714958 Review.
-
Oncogenic functions of the EMT-related transcription factor ZEB1 in breast cancer.J Transl Med. 2020 Feb 3;18(1):51. doi: 10.1186/s12967-020-02240-z. J Transl Med. 2020. PMID: 32014049 Free PMC article. Review.
Cited by
-
ELF3 is an antagonist of oncogenic-signalling-induced expression of EMT-TF ZEB1.Cancer Biol Ther. 2019;20(1):90-100. doi: 10.1080/15384047.2018.1507256. Epub 2018 Aug 27. Cancer Biol Ther. 2019. PMID: 30148686 Free PMC article.
-
Transcriptomic-Based Quantification of the Epithelial-Hybrid-Mesenchymal Spectrum across Biological Contexts.Biomolecules. 2021 Dec 25;12(1):29. doi: 10.3390/biom12010029. Biomolecules. 2021. PMID: 35053177 Free PMC article.
-
Mesenchymal to amoeboid transition is associated with stem-like features of melanoma cells.Cell Commun Signal. 2014 Apr 1;12:24. doi: 10.1186/1478-811X-12-24. Cell Commun Signal. 2014. PMID: 24690323 Free PMC article.
-
STAT3-Mediated Metabolic Reprograming in Cellular Transformation and Implications for Drug Resistance.Front Oncol. 2015 Jun 8;5:121. doi: 10.3389/fonc.2015.00121. eCollection 2015. Front Oncol. 2015. PMID: 26106584 Free PMC article. Review.
-
DNA Methylation Profiling of Breast Cancer Cell Lines along the Epithelial Mesenchymal Spectrum-Implications for the Choice of Circulating Tumour DNA Methylation Markers.Int J Mol Sci. 2018 Aug 28;19(9):2553. doi: 10.3390/ijms19092553. Int J Mol Sci. 2018. PMID: 30154364 Free PMC article.
References
-
- Tiwari N, Gheldof A, Tatari M, Christofori G. EMT as the ultimate survival mechanism of cancer cells. Semin Cancer Biol. 2012 - PubMed
-
- Yilmaz M, Christofori G. EMT, the cytoskeleton, and cancer cell invasion. Cancer Metastasis Rev. 2009;28:15–33. - PubMed
-
- Thiery JP, Acloque H, Huang RY, Nieto MA. Epithelial-mesenchymal transitions in development and disease. Cell. 2009;139:871–90. - PubMed
-
- Guadamillas MC, Cerezo A, Del Pozo MA. Overcoming anoikis--pathways to anchorage-independent growth in cancer. J Cell Sci. 124:3189–97. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
