

Intestinal epithelial vitamin D receptor signaling inhibits experimental colitis
- PMID: 23945234
- PMCID: PMC3754241
- DOI: 10.1172/JCI65842
Intestinal epithelial vitamin D receptor signaling inhibits experimental colitis
Abstract
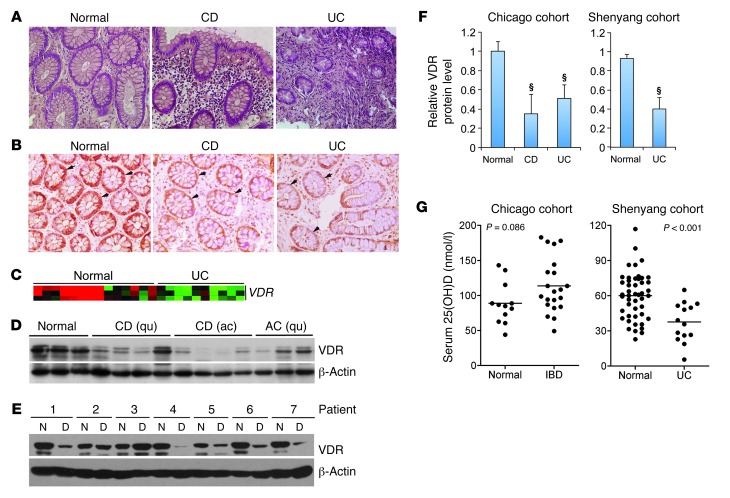
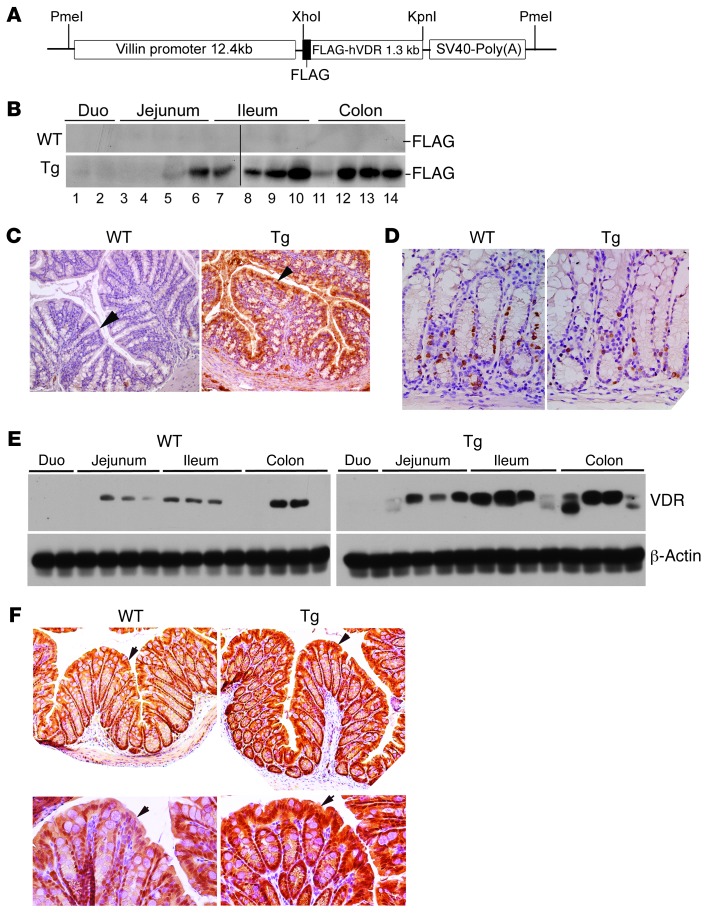
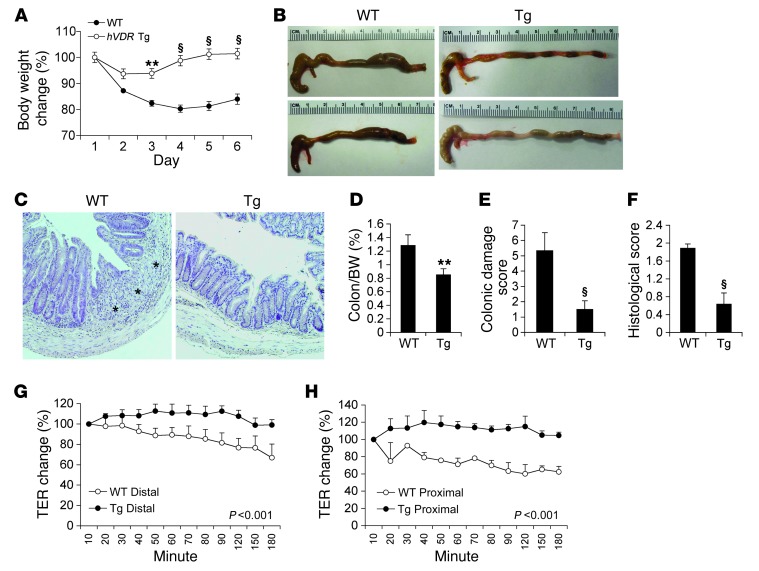
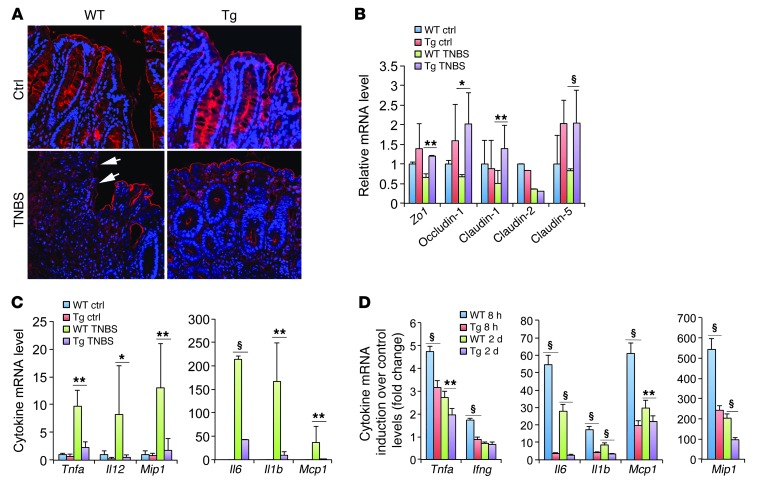
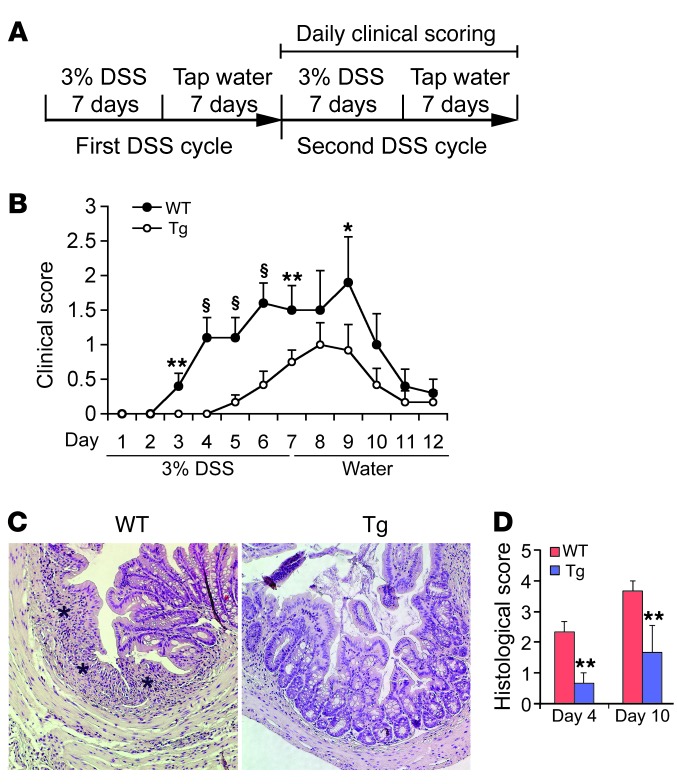
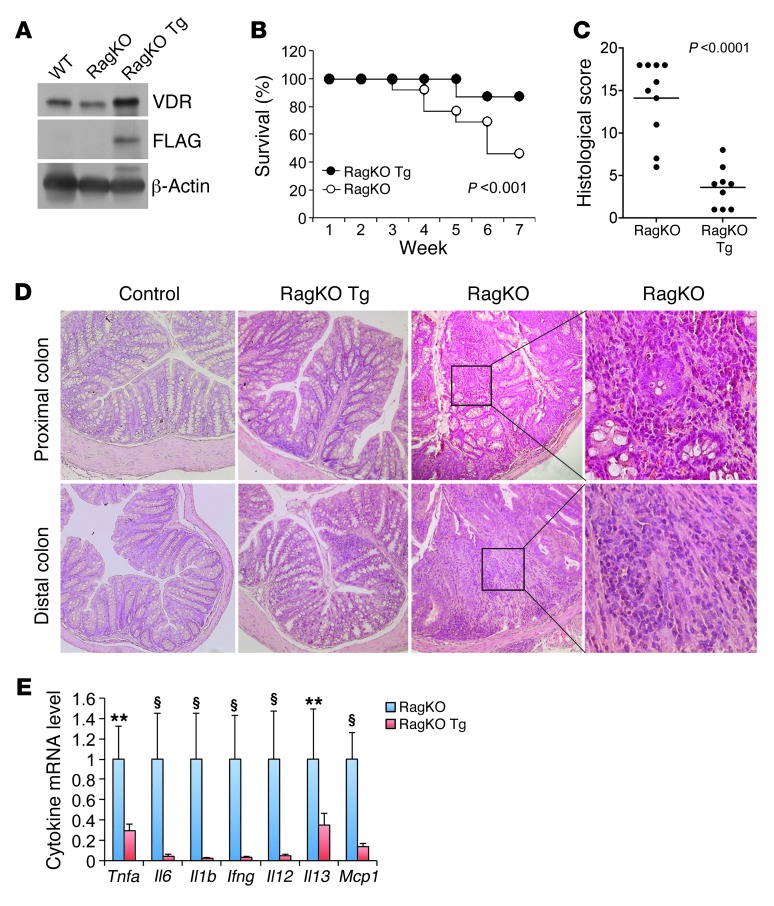
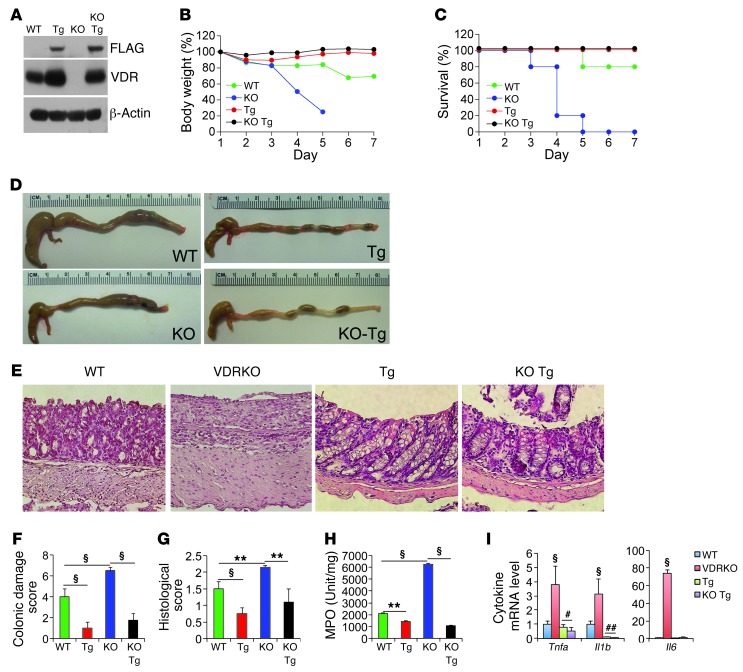
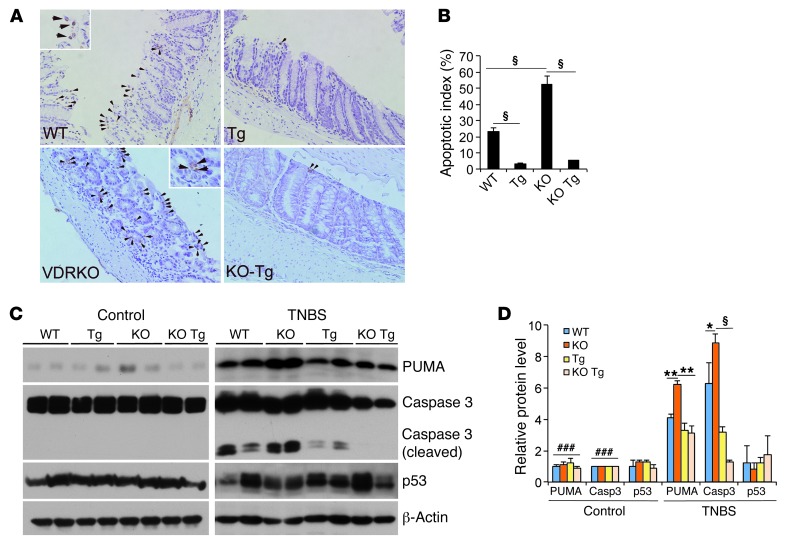
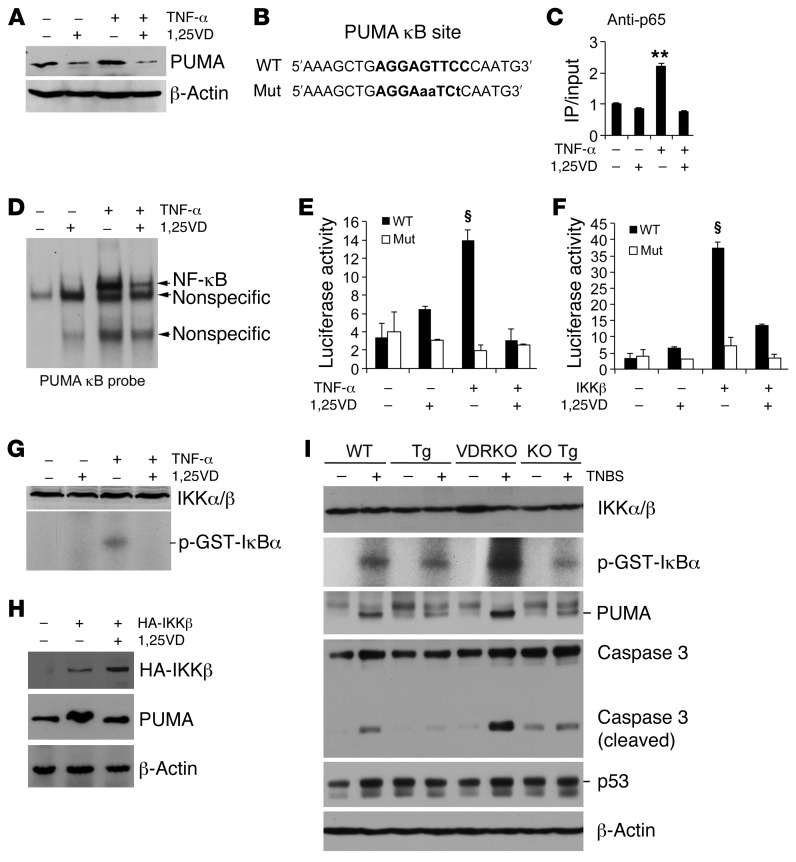
The inhibitory effects of vitamin D on colitis have been previously documented. Global vitamin D receptor (VDR) deletion exaggerates colitis, but the relative anticolitic contribution of epithelial and nonepithelial VDR signaling is unknown. Here, we showed that colonic epithelial VDR expression was substantially reduced in patients with Crohn's disease or ulcerative colitis. Moreover, targeted expression of human VDR (hVDR) in intestinal epithelial cells (IECs) protected mice from developing colitis. In experimental colitis models induced by 2,4,6-trinitrobenzenesulfonic acid, dextran sulfate sodium, or CD4(+)CD45RB(hi) T cell transfer, transgenic mice expressing hVDR in IECs were highly resistant to colitis, as manifested by marked reductions in clinical colitis scores, colonic histological damage, and colonic inflammation compared with WT mice. Reconstitution of Vdr-deficient IECs with the hVDR transgene completely rescued Vdr-null mice from severe colitis and death, even though the mice still maintained a hyperresponsive Vdr-deficient immune system. Mechanistically, VDR signaling attenuated PUMA induction in IECs by blocking NF-κB activation, leading to a reduction in IEC apoptosis. Together, these results demonstrate that gut epithelial VDR signaling inhibits colitis by protecting the mucosal epithelial barrier, and this anticolitic activity is independent of nonepithelial immune VDR actions.
Figures
References
-
- Fasano A, Shea-Donohue T. Mechanisms of disease: the role of intestinal barrier function in the pathogenesis of gastrointestinal autoimmune diseases. Nat Clin Pract Gastroenterol Hepatol. 2005;2(9):416–422. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
