

Place field repetition and purely local remapping in a multicompartment environment
- PMID: 23945240
- PMCID: PMC4400414
- DOI: 10.1093/cercor/bht198
Place field repetition and purely local remapping in a multicompartment environment
Abstract
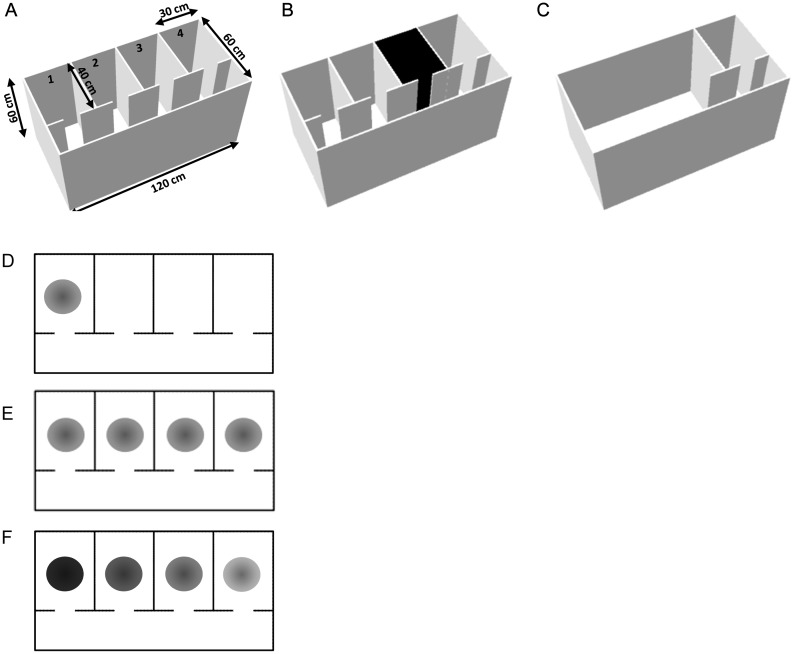
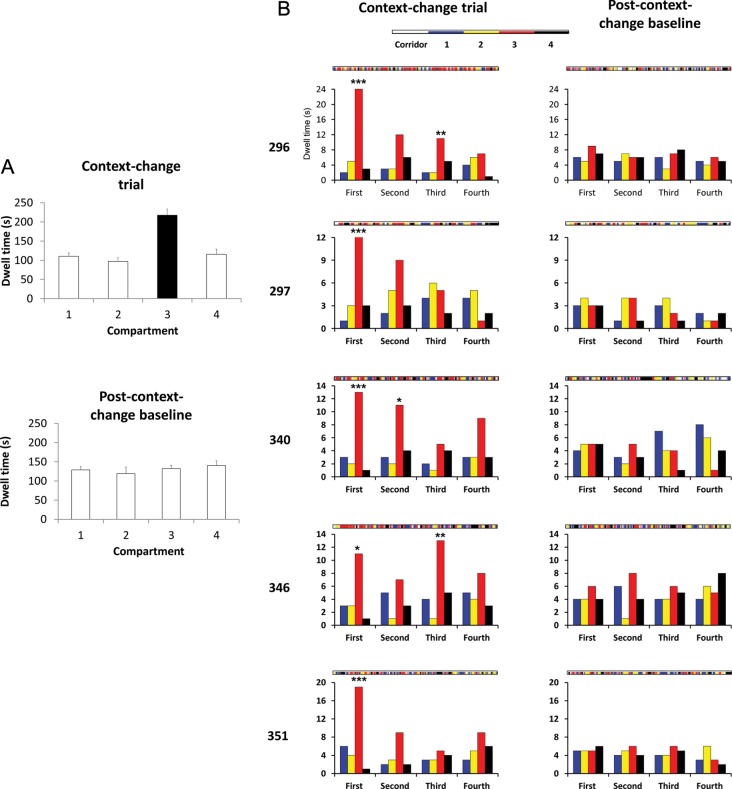
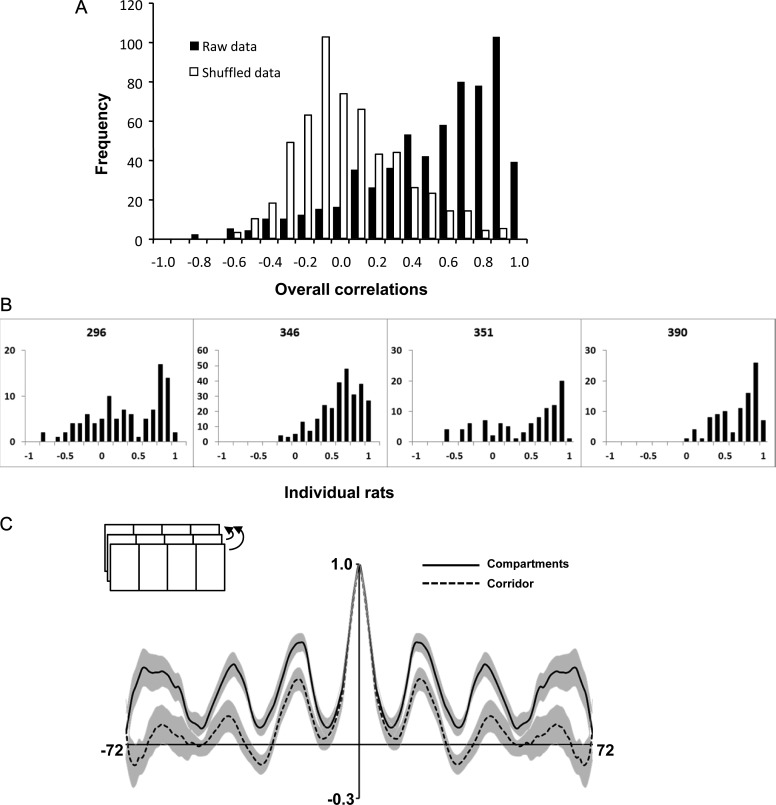
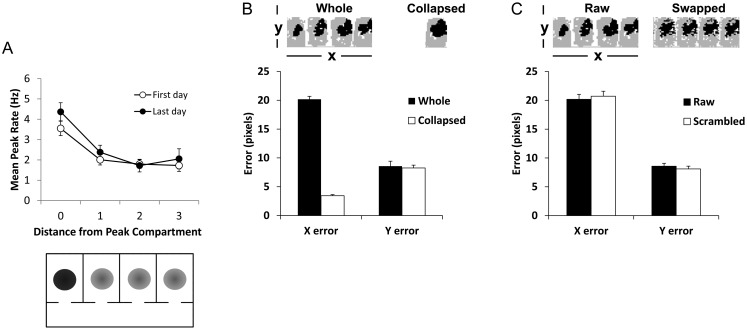
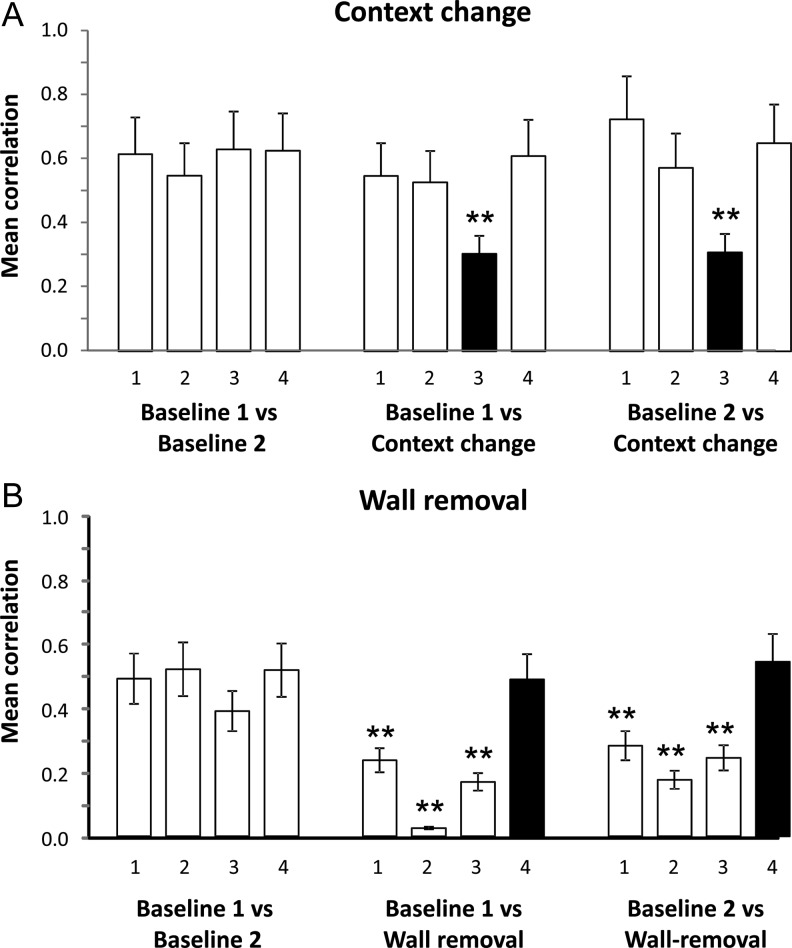
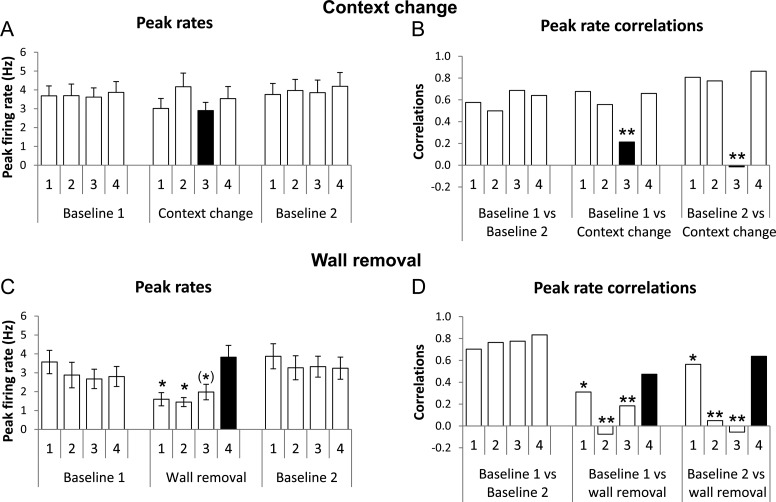
Hippocampal place cells support spatial memory using sensory information from the environment and self-motion information to localize their firing fields. Currently, there is disagreement about whether CA1 place cells can use pure self-motion information to disambiguate different compartments in environments containing multiple visually identical compartments. Some studies report that place cells can disambiguate different compartments, while others report that they do not. Furthermore, while numerous studies have examined remapping, there has been little examination of remapping in different subregions of a single environment. Is remapping purely local or do place fields in neighboring, unaffected, regions detect the change? We recorded place cells as rats foraged across a 4-compartment environment and report 3 new findings. First, we find that, unlike studies in which rats foraged in 2 compartments, place fields showed a high degree of spatial repetition with a slight degree of rate-based discrimination. Second, this repetition does not diminish with extended experience. Third, remapping was found to be purely local for both geometric change and contextual change. Our results reveal the limited capacity of the path integrator to drive pattern separation in hippocampal representations, and suggest that doorways may play a privileged role in segmenting the neural representation of space.
Keywords: grid cells; path integration; place cells; rat; spatial memory.
© The Author 2013. Published by Oxford University Press.
Figures



References
-
- Boccara CN, Sargolini F, Thoresen VH, Solstad T, Witter MP, Moser EI, Moser M-B. Grid cells in pre-and parasubiculum. Nat Neurosci. 2010;13:987–994. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
