

Rescue of peripheral and CNS axon defects in mice lacking NMNAT2
- PMID: 23946398
- PMCID: PMC6705152
- DOI: 10.1523/JNEUROSCI.1534-13.2013
Rescue of peripheral and CNS axon defects in mice lacking NMNAT2
Abstract
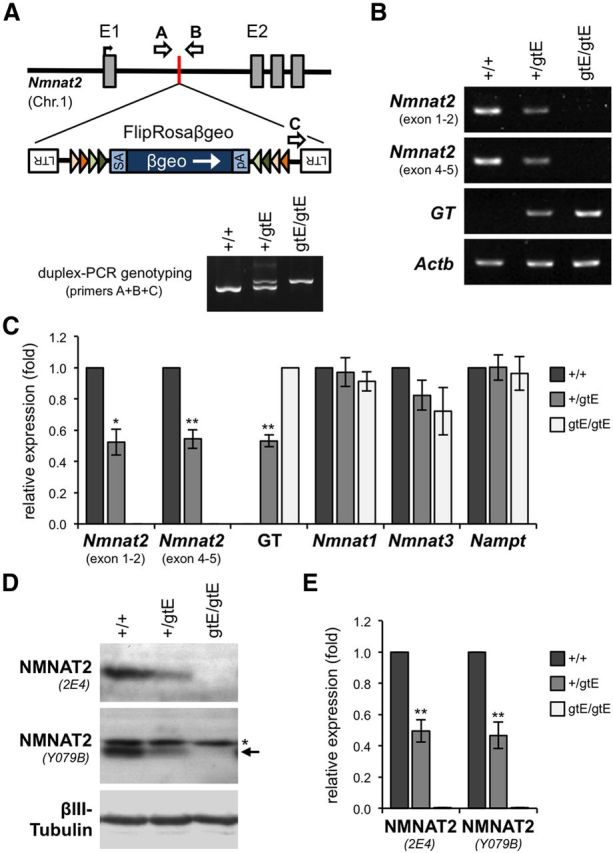
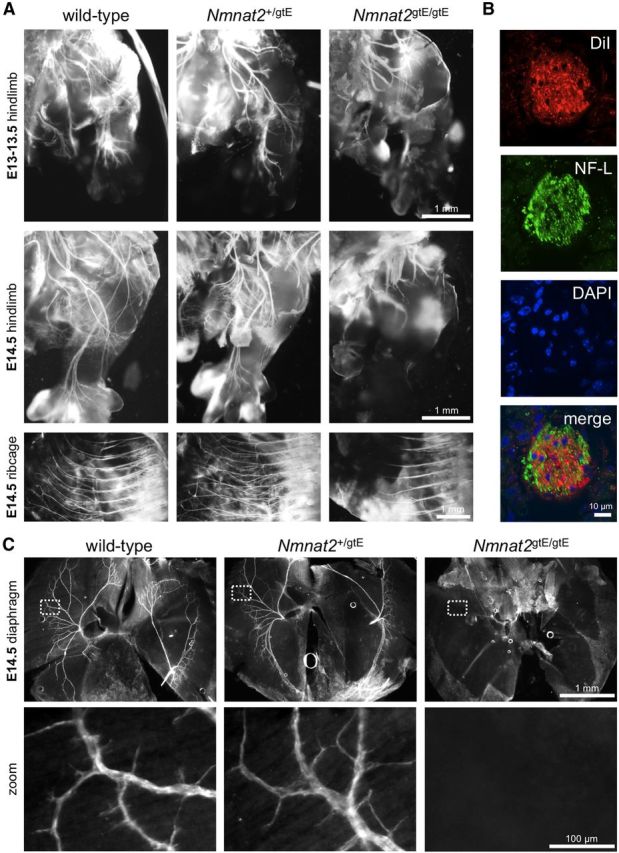
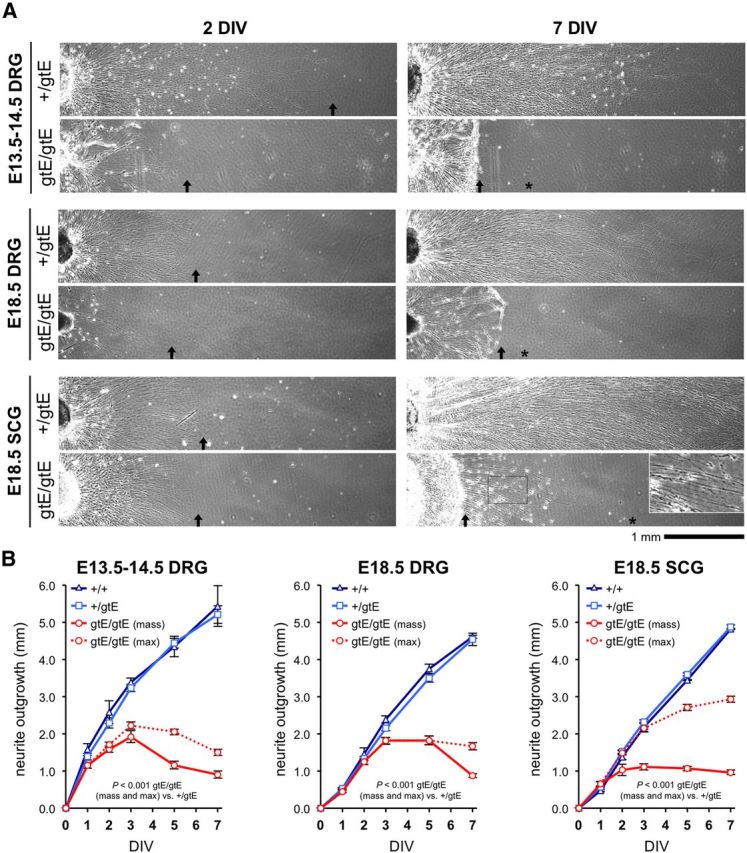
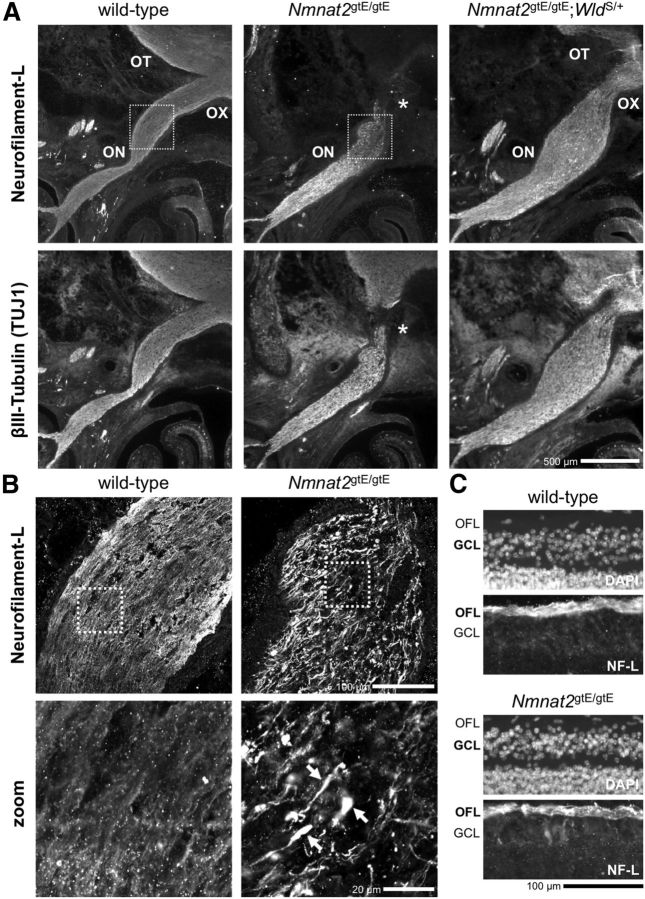
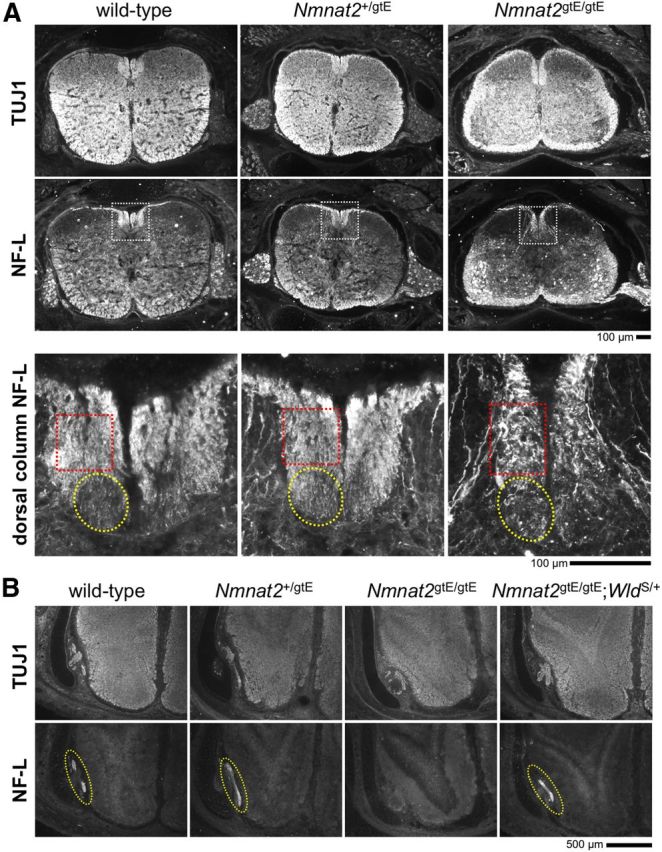
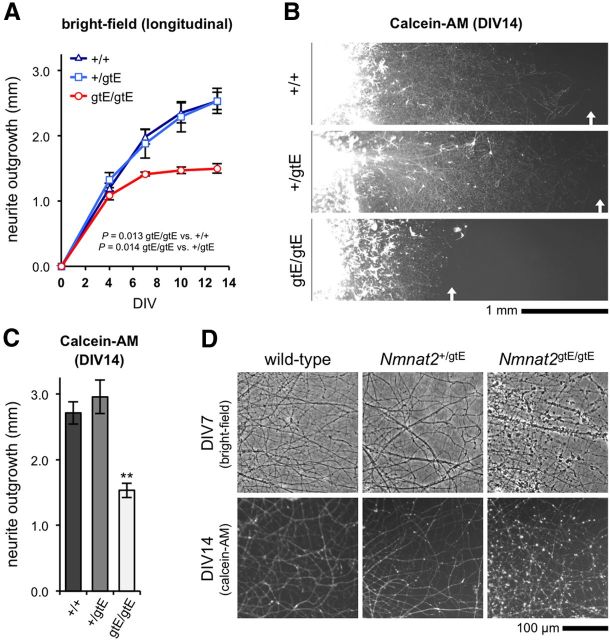
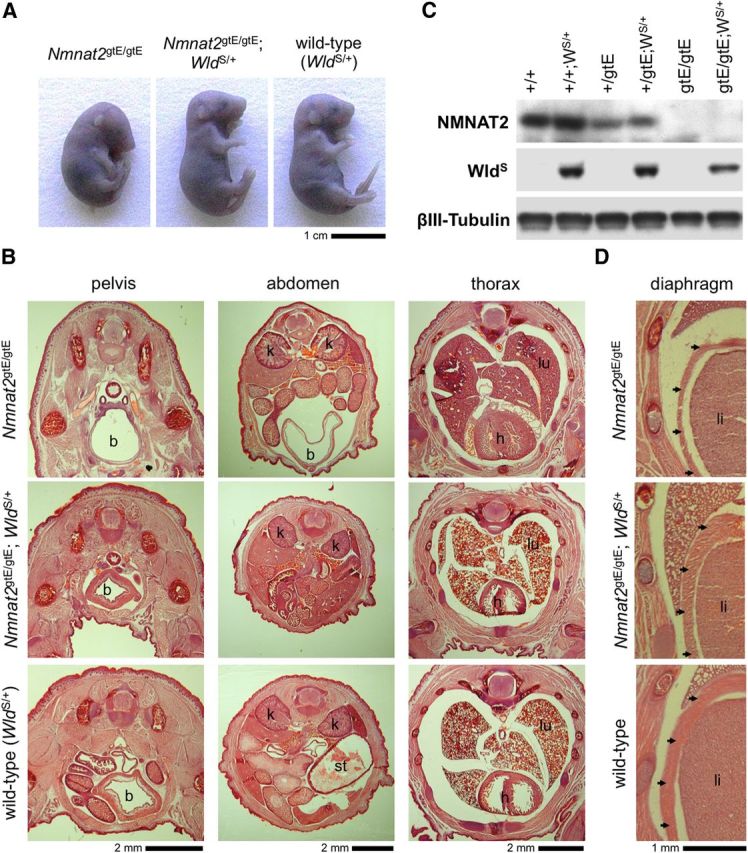
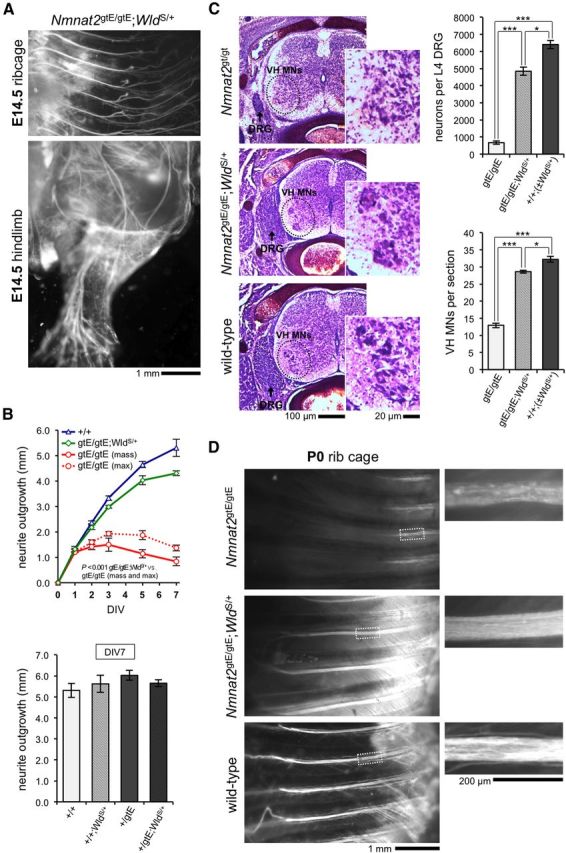
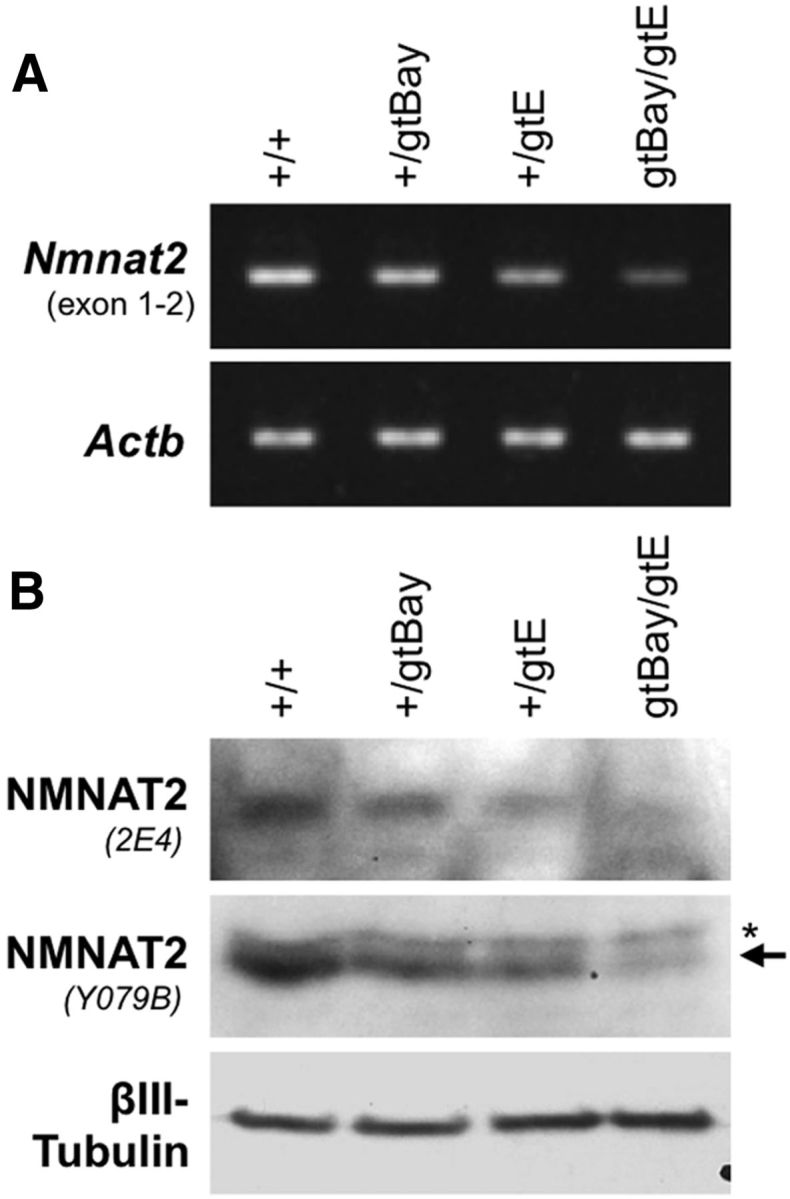
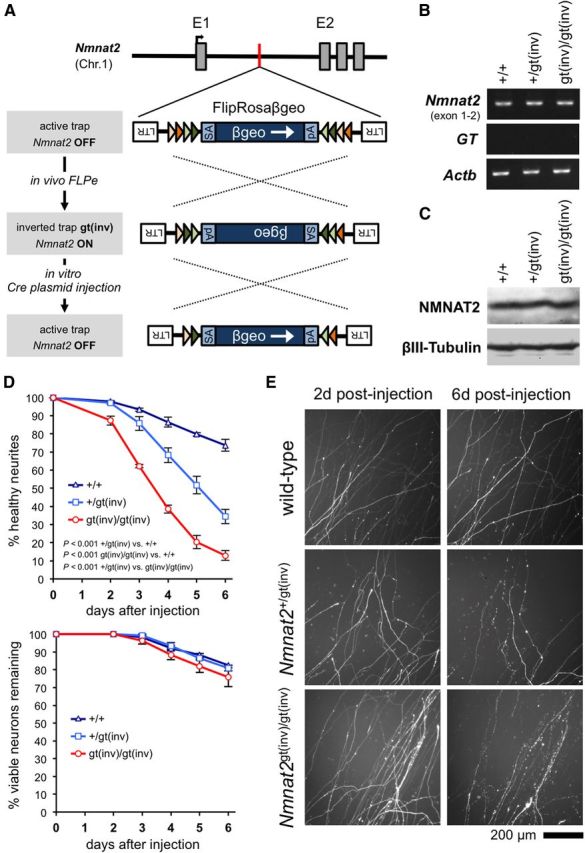
NMNAT2 is an NAD(+)-synthesizing enzyme with an essential axon maintenance role in primary culture neurons. We have generated an Nmnat2 gene trap mouse to examine the role of NMNAT2 in vivo. Homozygotes die perinatally with a severe peripheral nerve/axon defect and truncated axons in the optic nerve and other CNS regions. The cause appears to be limited axon extension, rather than dying-back degeneration of existing axons, which was previously proposed for the NMNAT2-deficient Blad mutant mouse. Neurite outgrowth in both PNS and CNS neuronal cultures consistently stalls at 1-2 mm, similar to the length of truncated axons in the embryos. Crucially, this suggests an essential role for NMNAT2 during axon growth. In addition, we show that the Wallerian degeneration slow protein (Wld(S)), a more stable, aberrant NMNAT that can substitute the axon maintenance function of NMNAT2 in primary cultures, can also correct developmental defects associated with NMNAT2 deficiency. This is dose-dependent, with extension of life span to at least 3 months by homozygous levels of Wld(S) the most obvious manifestation. Finally, we propose that endogenous mechanisms also compensate for otherwise limiting levels of NMNAT2. This could explain our finding that conditional silencing of a single Nmnat2 allele triggers substantial degeneration of established neurites, whereas similar, or greater, reduction of NMNAT2 in constitutively depleted neurons is compatible with normal axon growth and survival. A requirement for NMNAT2 for both axon growth and maintenance suggests that reduced levels could impair axon regeneration as well as axon survival in aging and disease.
Figures
Comment in
-
Axonal trafficking of NMNAT2 and its roles in axon growth and survival in vivo.Bioarchitecture. 2013 Sep-Dec;3(5):133-40. doi: 10.4161/bioa.27049. Epub 2013 Nov 7. Bioarchitecture. 2013. PMID: 24284888 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases