

FAD synthesis and degradation in the nucleus create a local flavin cofactor pool
- PMID: 23946482
- PMCID: PMC3790006
- DOI: 10.1074/jbc.M113.500066
FAD synthesis and degradation in the nucleus create a local flavin cofactor pool
Abstract
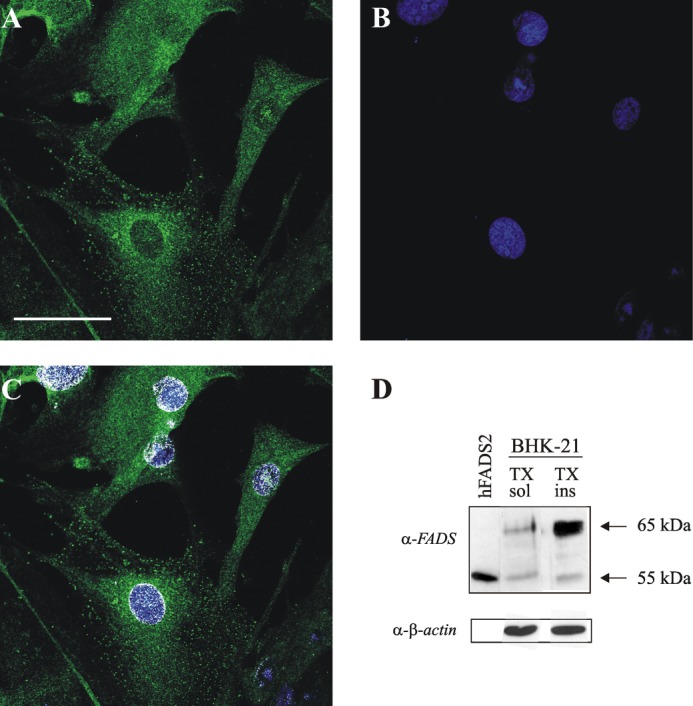
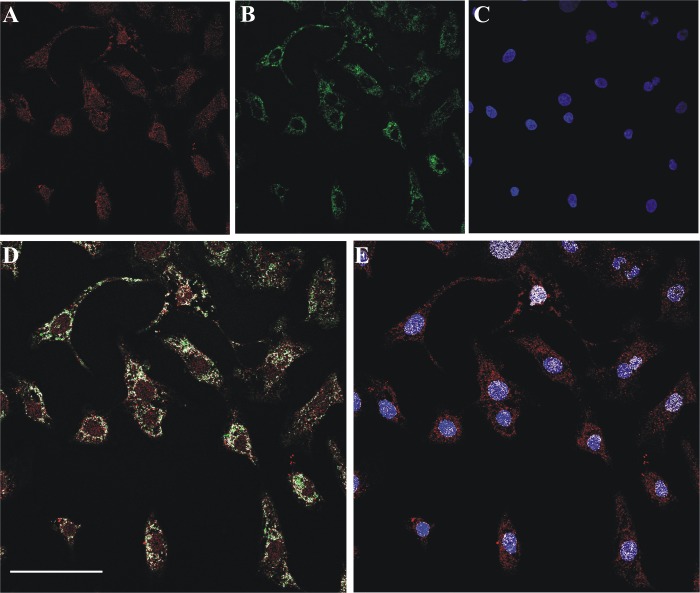
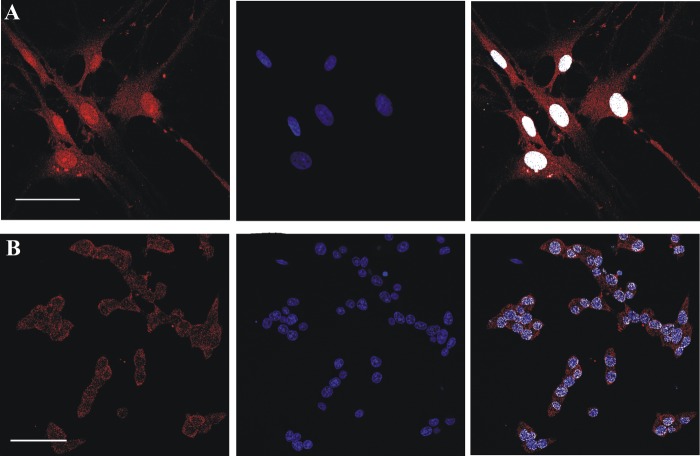
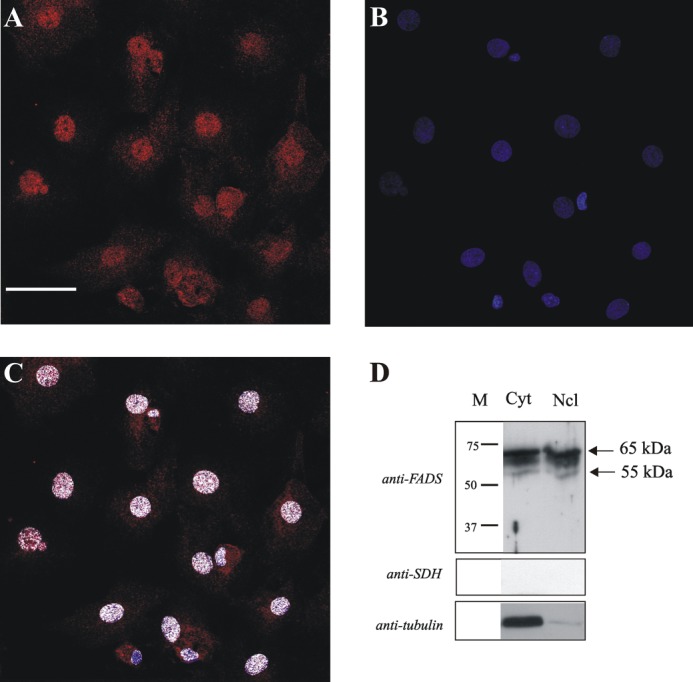
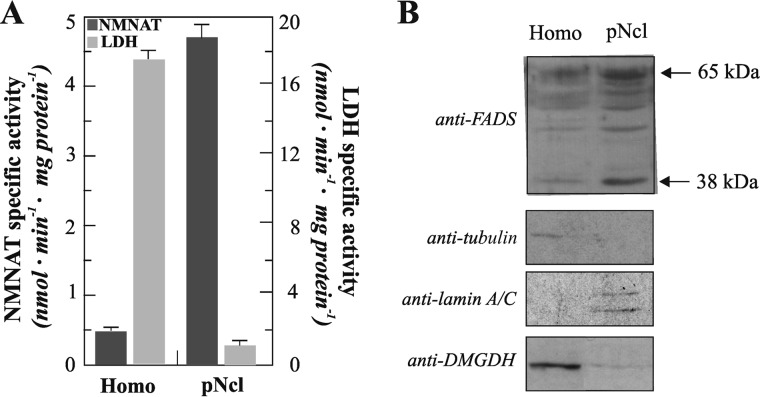
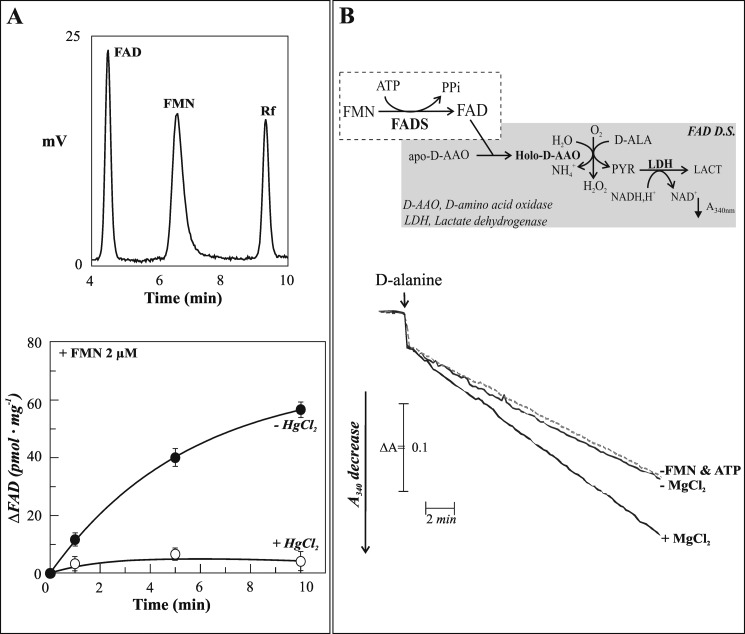
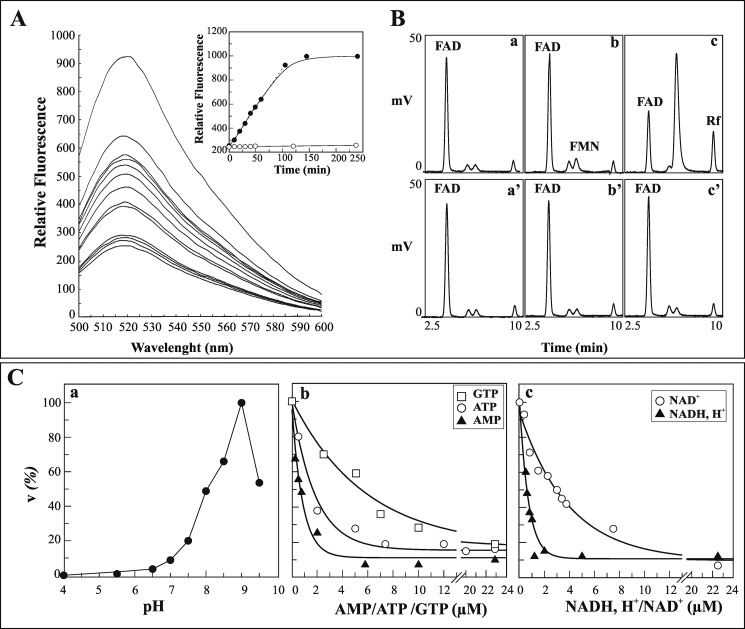
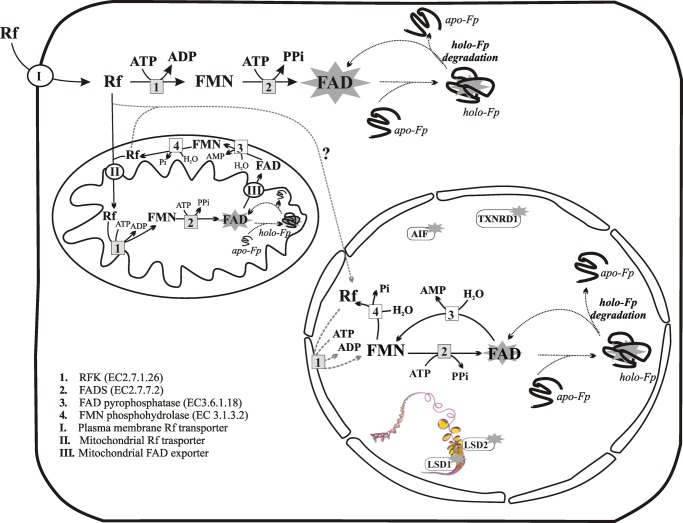
FAD is a redox cofactor ensuring the activity of many flavoenzymes mainly located in mitochondria but also relevant for nuclear redox activities. The last enzyme in the metabolic pathway producing FAD is FAD synthase (EC 2.7.7.2), a protein known to be localized both in cytosol and in mitochondria. FAD degradation to riboflavin occurs via still poorly characterized enzymes, possibly belonging to the NUDIX hydrolase family. By confocal microscopy and immunoblotting experiments, we demonstrate here the existence of FAD synthase in the nucleus of different experimental rat models. HPLC experiments demonstrated that isolated rat liver nuclei contain ∼300 pmol of FAD·mg(-1) protein, which was mainly protein-bound FAD. A mean FAD synthesis rate of 18.1 pmol·min(-1)·mg(-1) protein was estimated by both HPLC and continuous coupled enzymatic spectrophotometric assays. Rat liver nuclei were also shown to be endowed with a FAD pyrophosphatase that hydrolyzes FAD with an optimum at alkaline pH and is significantly inhibited by adenylate-containing nucleotides. The coordinate activity of these FAD forming and degrading enzymes provides a potential mechanism by which a dynamic pool of flavin cofactor is created in the nucleus. These data, which significantly add to the biochemical comprehension of flavin metabolism and its subcellular compartmentation, may also provide the basis for a more detailed comprehension of the role of flavin homeostasis in biologically and clinically relevant epigenetic events.
Keywords: Confocal Microscopy; FAD; FAD Hydrolysis; FAD Synthase; FMN; NUDIX Hydrolase; Nucleus; Rat; Riboflavin.
Figures
References
-
- Massey V. (1995) Introduction. Flavoprotein structure and mechanism. FASEB J. 9, 473–475 - PubMed
-
- Massey V. (2000) The chemical and biological versatility of riboflavin. Biochem. Soc. Trans. 28, 283–296 - PubMed
-
- McCormick D. B. (1989) Two interconnected B vitamins. Riboflavin and pyridoxine. Physiol. Rev. 69, 1170–1198 - PubMed
-
- Macheroux P., Kappes B., Ealick S. E. (2011) Flavogenomics. A genomic and structural view of flavin-dependent proteins. FEBS J. 278, 2625–2634 - PubMed
-
- Joosten V., van Berkel W. J. (2007) Flavoenzymes. Curr. Opin. Chem. Biol. 11, 195–202 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
