

Exo70 generates membrane curvature for morphogenesis and cell migration
- PMID: 23948253
- PMCID: PMC3762978
- DOI: 10.1016/j.devcel.2013.07.007
Exo70 generates membrane curvature for morphogenesis and cell migration
Abstract
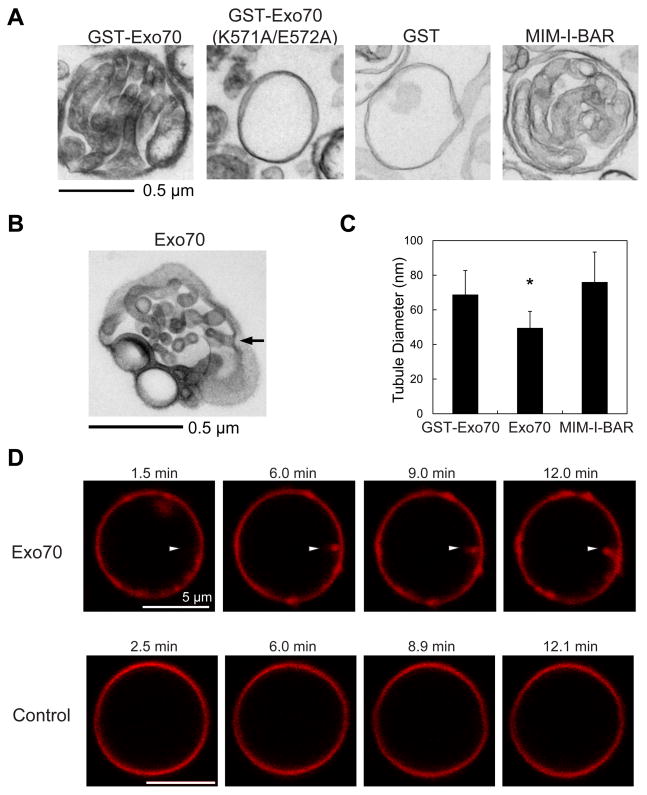
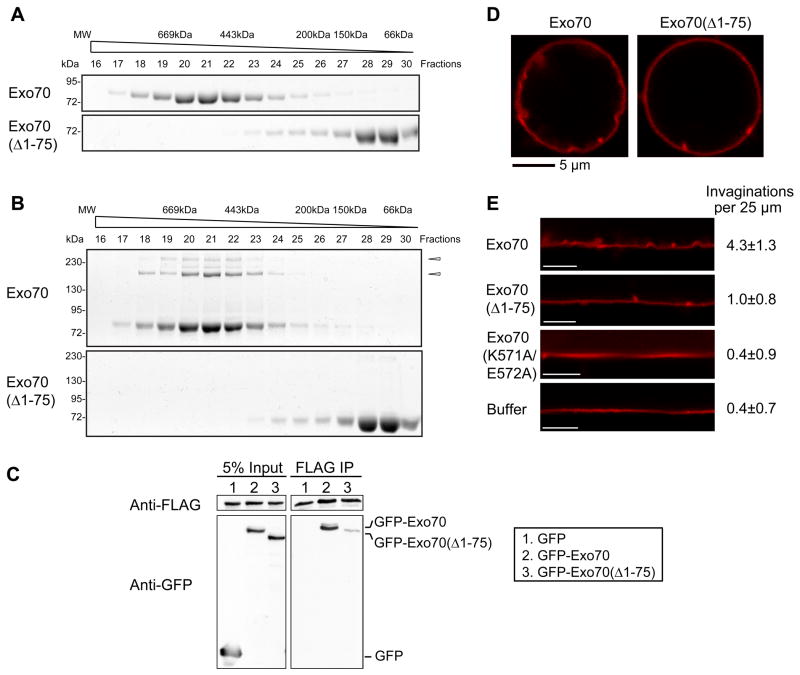
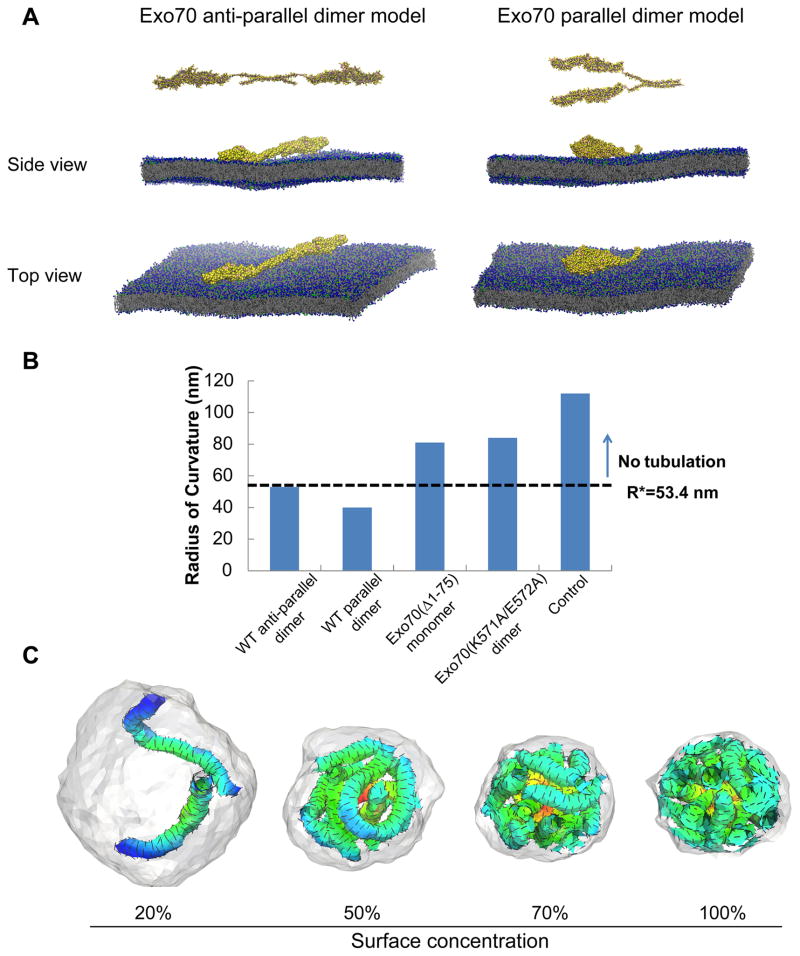
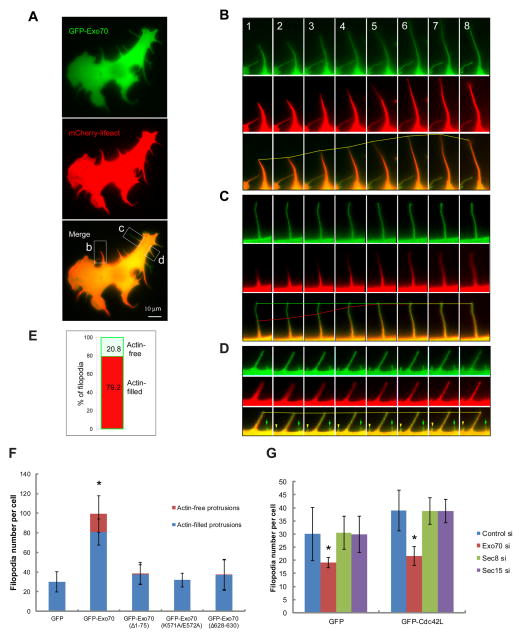
Dynamic shape changes of the plasma membrane are fundamental to many processes, ranging from morphogenesis and cell migration to phagocytosis and viral propagation. Here, we demonstrate that Exo70, a component of the exocyst complex, induces tubular membrane invaginations toward the lumen of synthetic vesicles in vitro and generates protrusions on the surface of cells. Biochemical analyses using Exo70 mutants and independent molecular dynamics simulations based on Exo70 structure demonstrate that Exo70 generates negative membrane curvature through an oligomerization-based mechanism. In cells, the membrane-deformation function of Exo70 is required for protrusion formation and directional cell migration. Exo70 thus represents a membrane-bending protein that may couple actin dynamics and plasma membrane remodeling for morphogenesis.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
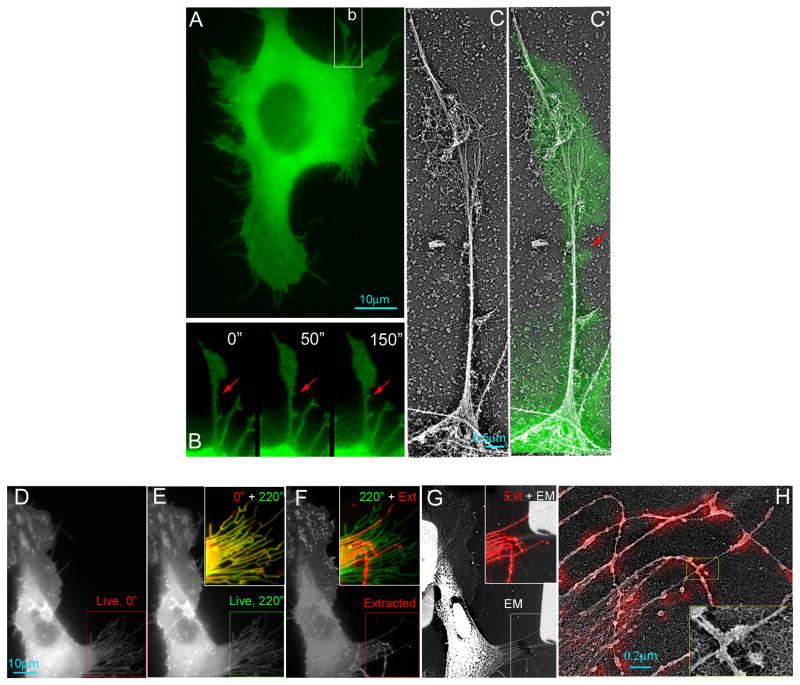
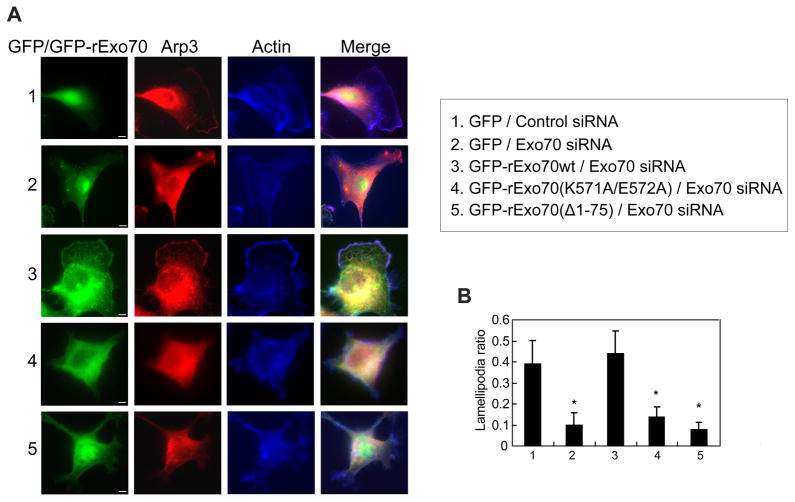
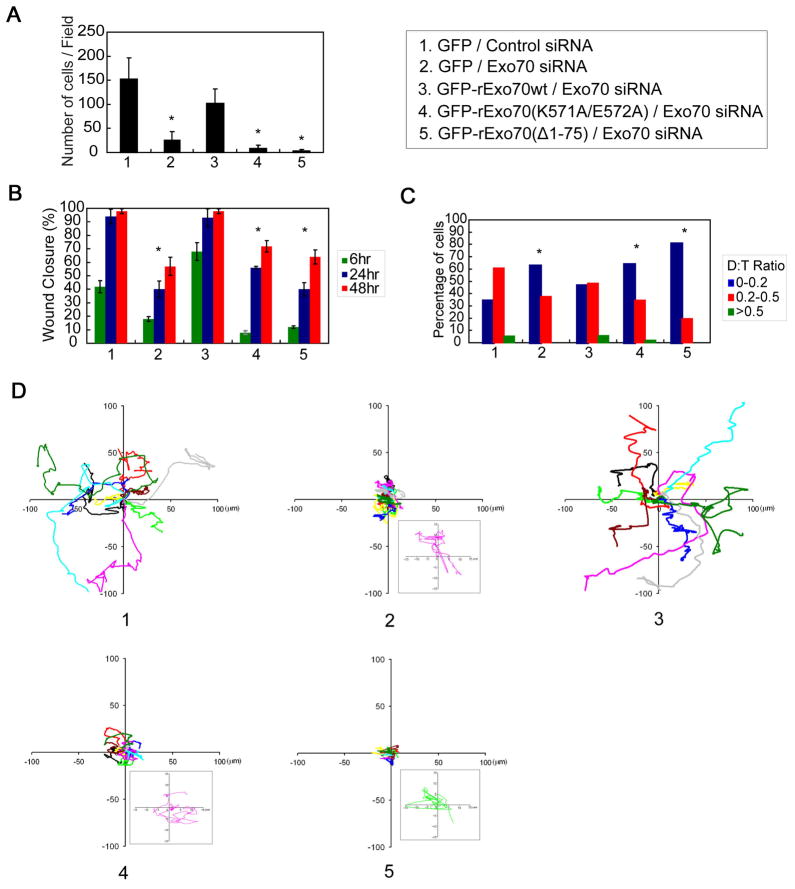
References
-
- Angelova MI, Dimitrov DS. Liposome electroformation. Faraday Discuss Chem Soc. 1986;81:303–311.
-
- Bereiter-Hahn J. Mechanics of crawling cells. Med Eng Phys. 2005;27:743–753. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
