

New genes as drivers of phenotypic evolution
- PMID: 23949544
- PMCID: PMC4236023
- DOI: 10.1038/nrg3521
New genes as drivers of phenotypic evolution
Erratum in
- Nat Rev Genet. 2013 Oct;14(10):744
Abstract
During the course of evolution, genomes acquire novel genetic elements as sources of functional and phenotypic diversity, including new genes that originated in recent evolution. In the past few years, substantial progress has been made in understanding the evolution and phenotypic effects of new genes. In particular, an emerging picture is that new genes, despite being present in the genomes of only a subset of species, can rapidly evolve indispensable roles in fundamental biological processes, including development, reproduction, brain function and behaviour. The molecular underpinnings of how new genes can develop these roles are starting to be characterized. These recent discoveries yield fresh insights into our broad understanding of biological diversity at refined resolution.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
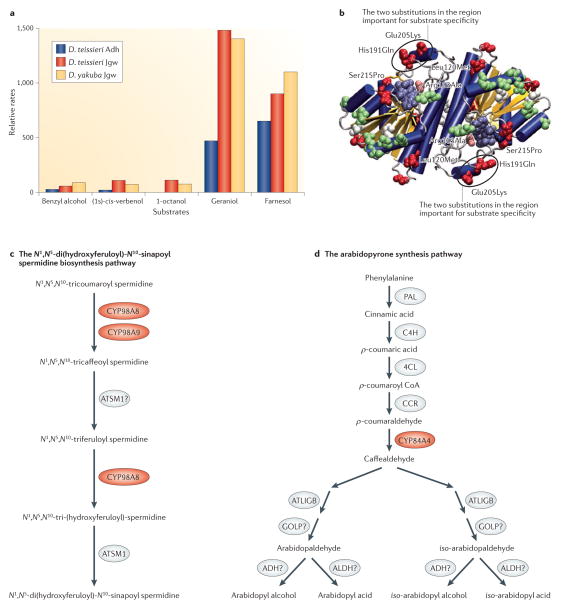
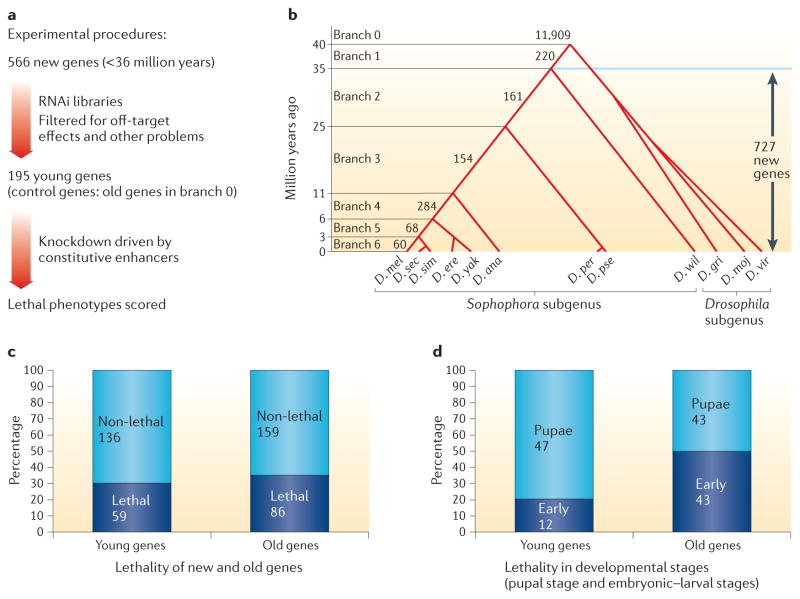
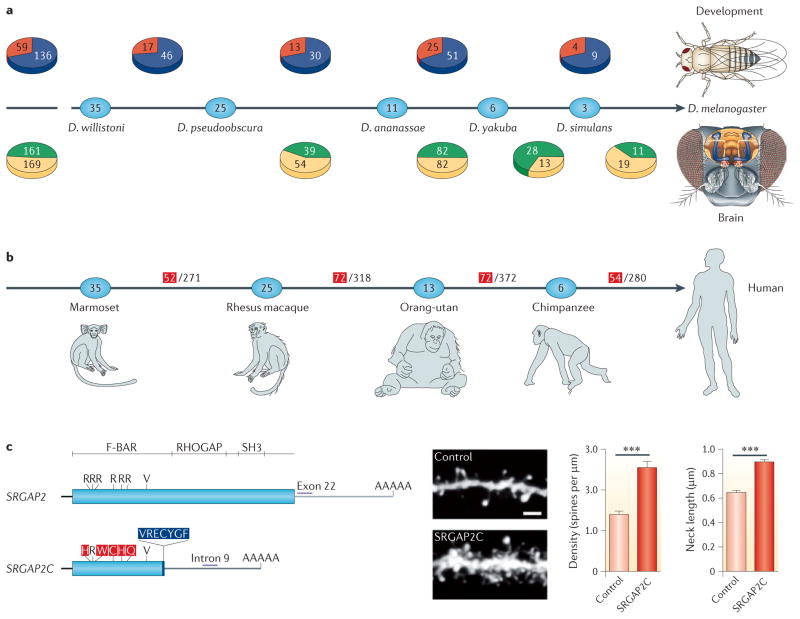
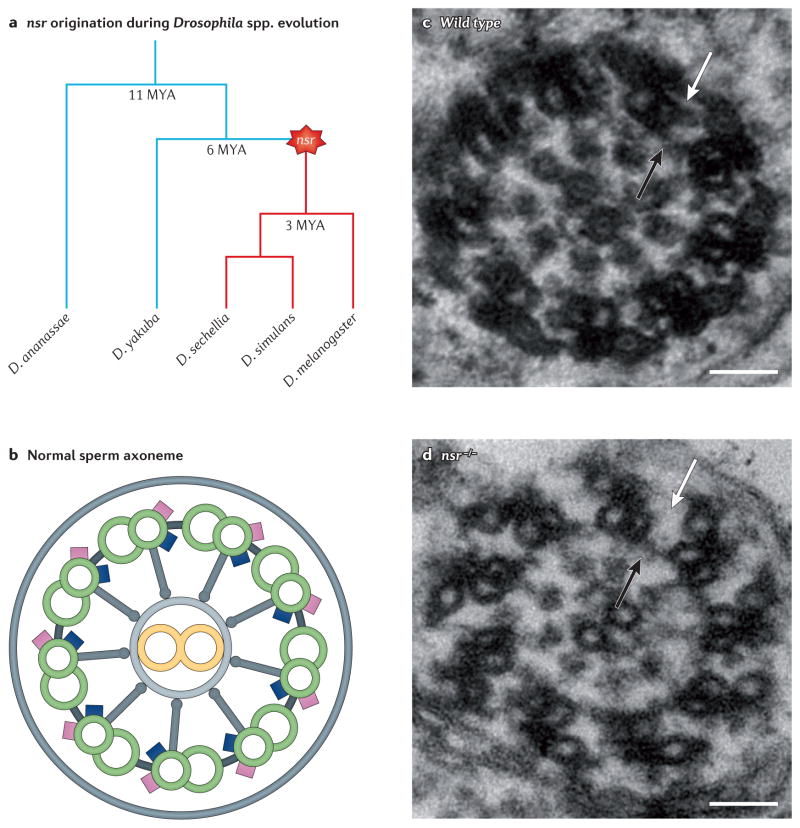
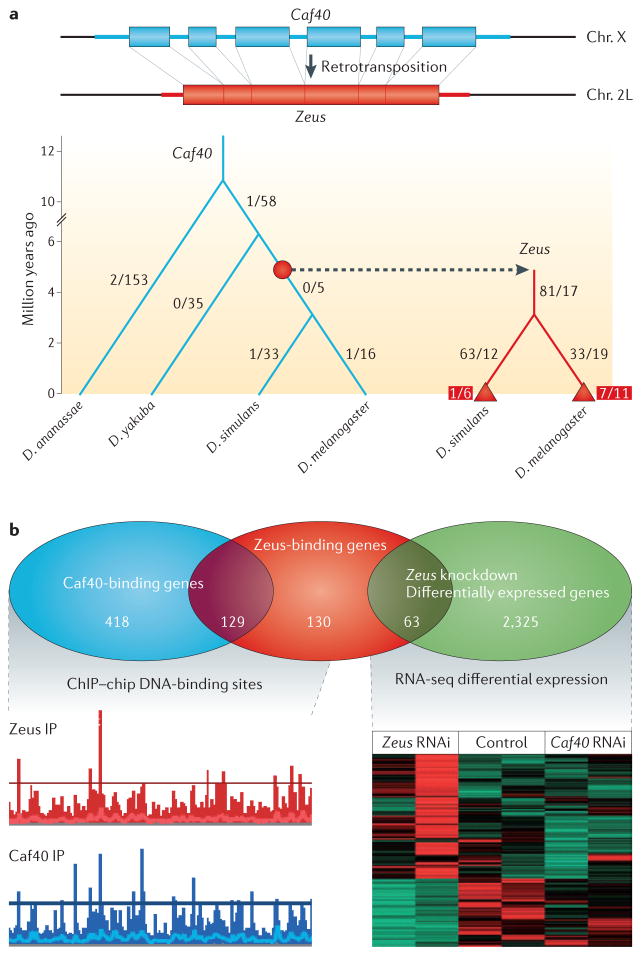
References
-
- Long M, Betran E, Thornton K, Wang W. The origin of new genes: glimpses from the young and old. Nature Rev Genet. 2003;4:865–875. - PubMed
-
- Ding Y, Zhou Q, Wang W. Origins of new genes and evolution of their novel functions. Annu Rev Ecol Evol Syst. 2012;43:345–363.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
