

Effect of temperature on photosynthesis and growth in marine Synechococcus spp
- PMID: 23950220
- PMCID: PMC3793060
- DOI: 10.1104/pp.113.221937
Effect of temperature on photosynthesis and growth in marine Synechococcus spp
Abstract
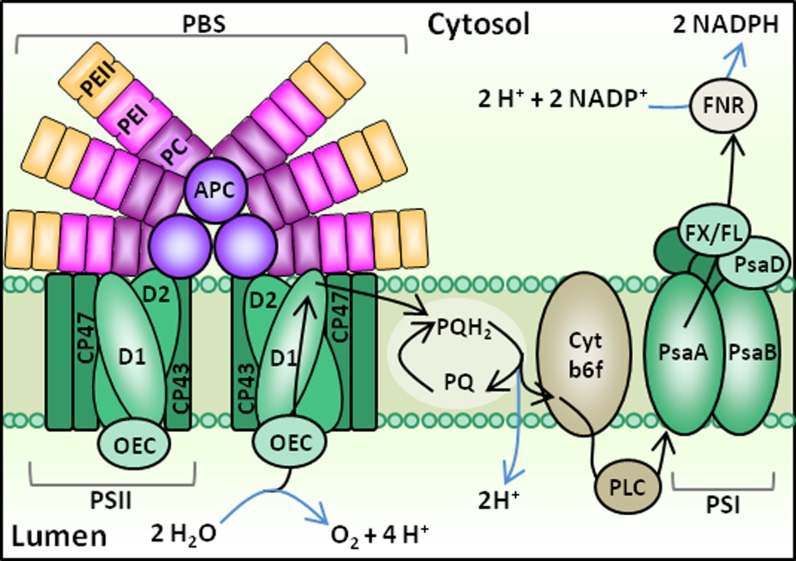
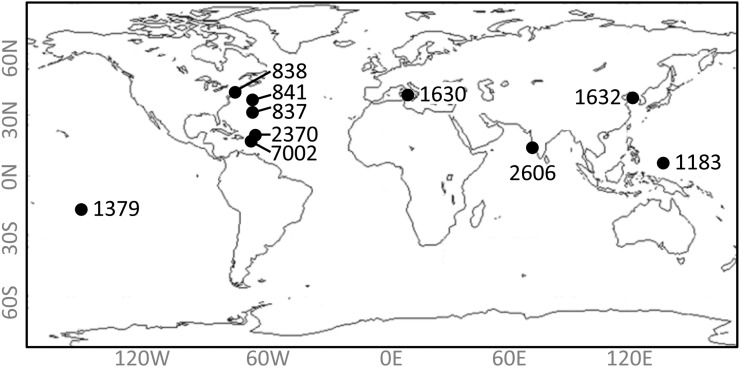
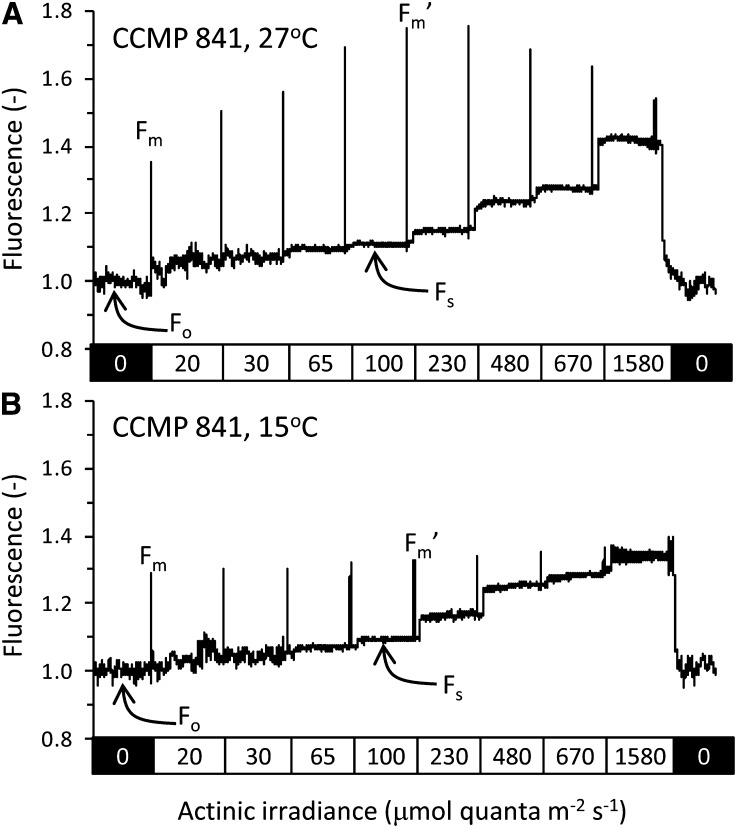
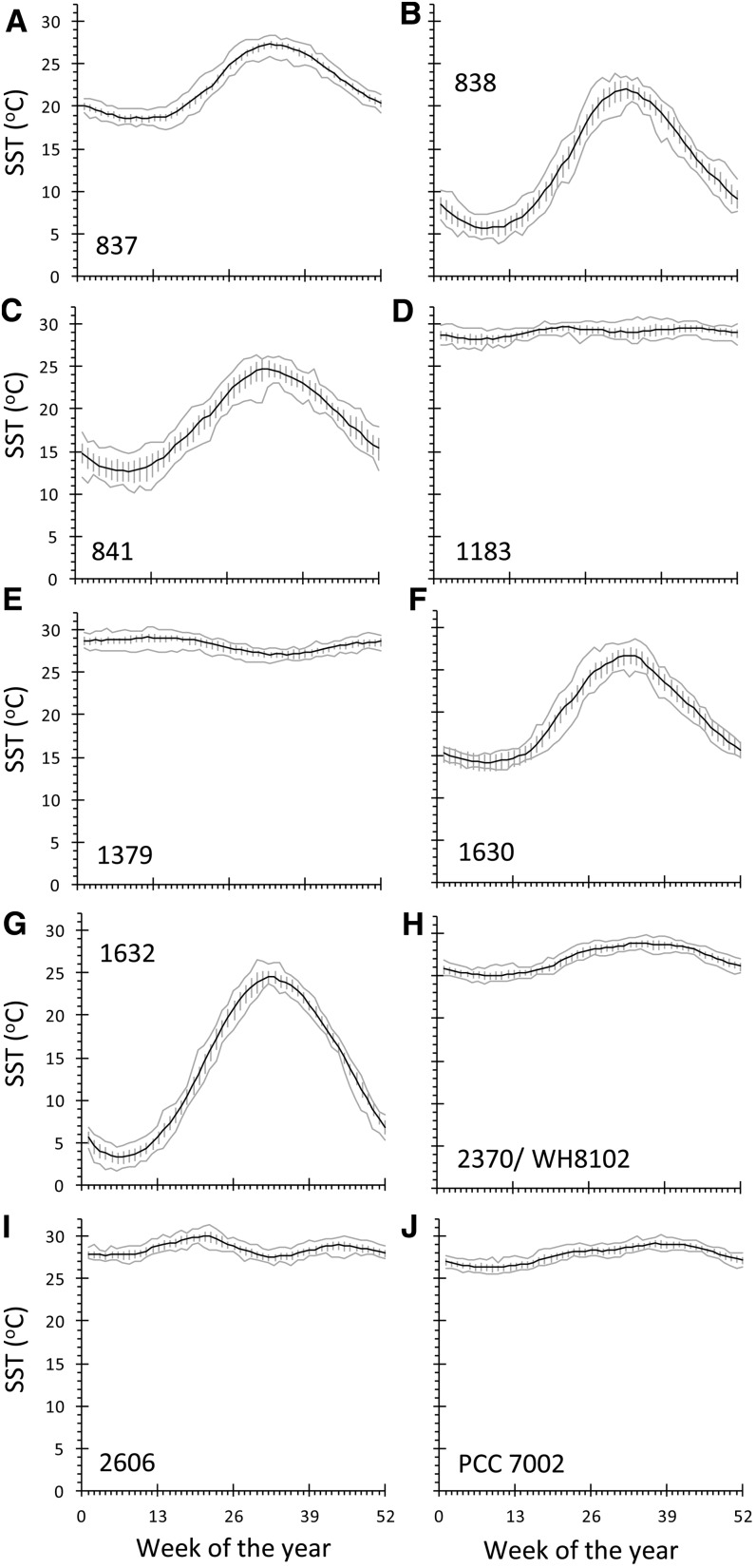
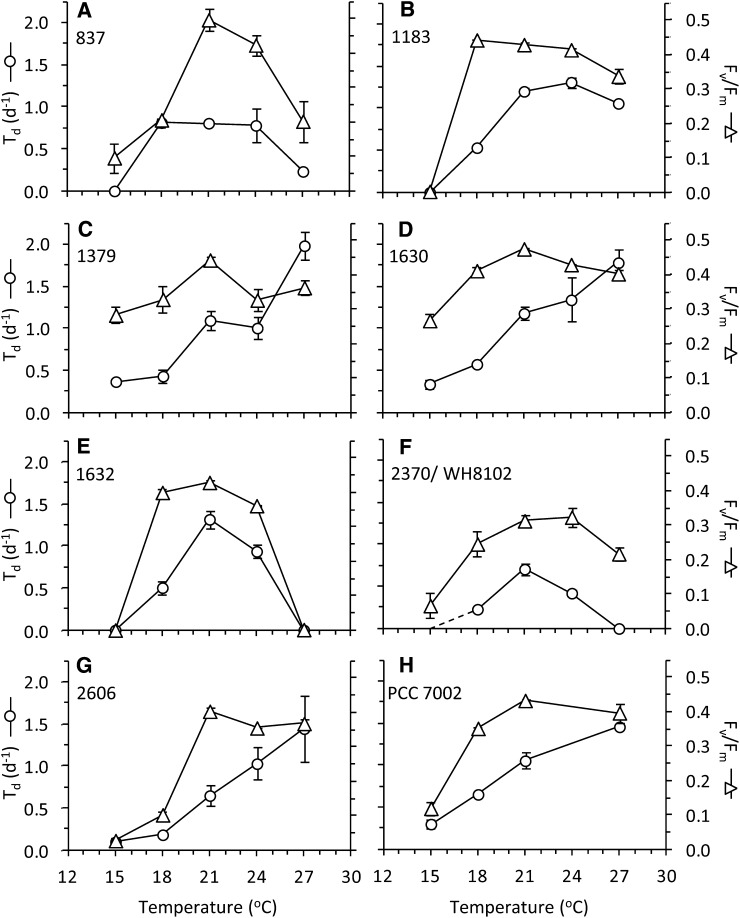
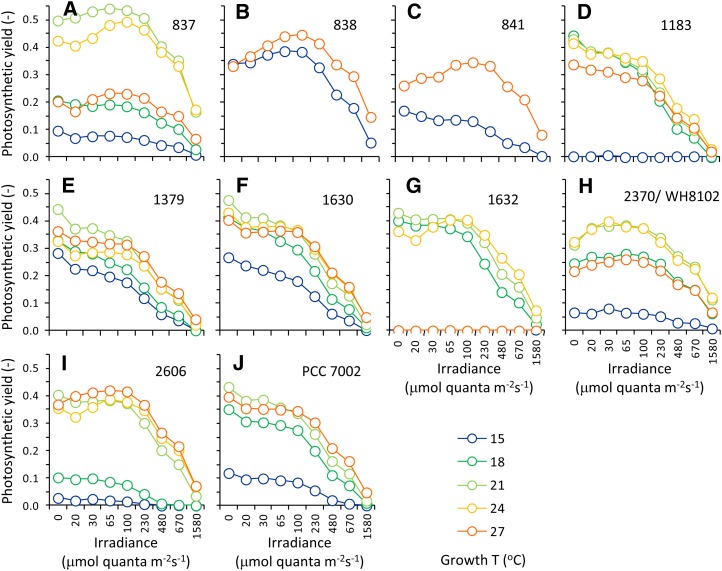
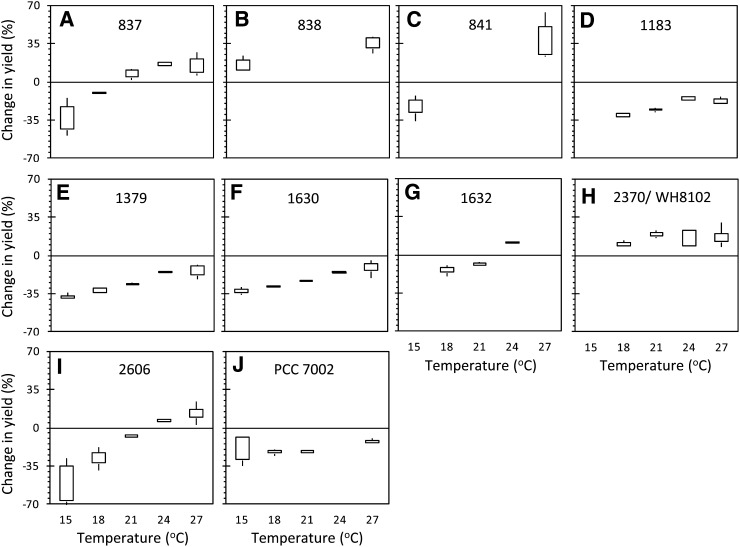
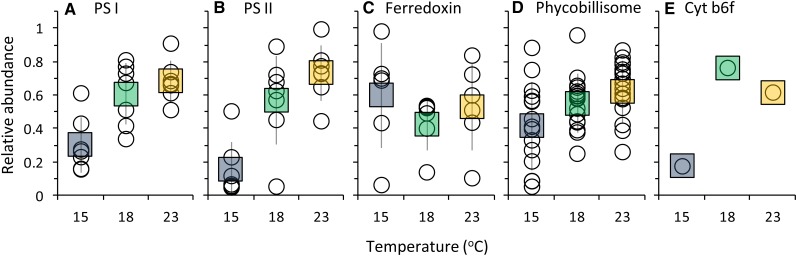
In this study, we develop a mechanistic understanding of how temperature affects growth and photosynthesis in 10 geographically and physiologically diverse strains of Synechococcus spp. We found that Synechococcus spp. are able to regulate photochemistry over a range of temperatures by using state transitions and altering the abundance of photosynthetic proteins. These strategies minimize photosystem II (PSII) photodamage by keeping the photosynthetic electron transport chain (ETC), and hence PSII reaction centers, more oxidized. At temperatures that approach the optimal growth temperature of each strain when cellular demand for reduced nicotinamide adenine dinucleotide phosphate (NADPH) is greatest, the phycobilisome (PBS) antenna associates with PSII, increasing the flux of electrons into the ETC. By contrast, under low temperature, when slow growth lowers the demand for NADPH and linear ETC declines, the PBS associates with photosystem I. This favors oxidation of PSII and potential increase in cyclic electron flow. For Synechococcus sp. WH8102, growth at higher temperatures led to an increase in the abundance of PBS pigment proteins, as well as higher abundance of subunits of the PSII, photosystem I, and cytochrome b6f complexes. This would allow cells to increase photosynthetic electron flux to meet the metabolic requirement for NADPH during rapid growth. These PBS-based temperature acclimation strategies may underlie the larger geographic range of this group relative to Prochlorococcus spp., which lack a PBS.
Figures

References
-
- Agusti S. (2004) Viability and niche segregation of Prochlorococcus and Synechococcus cells across the Central Atlantic Ocean. Aquat Microb Ecol 36: 53–59
-
- Aro EM, Virgin I, Andersson B. (1993) Photoinhibition of photosystem II. Inactivation, protein damage, and turnover. Biochim Biophys Acta 1143: 113–134 - PubMed
-
- Bailey S, Grossman A. (2008) Photoprotection in cyanobacteria: regulation of light harvesting. Photochem Photobiol 84: 1410–1420 - PubMed
-
- Bailey S, Melis A, Mackey KR, Cardol P, Finazzi G, van Dijken G, Berg GM, Arrigo K, Shrager J, Grossman A. (2008) Alternative photosynthetic electron flow to oxygen in marine Synechococcus. Biochim Biophys Acta 1777: 269–276 - PubMed
-
- Bald D, Kruip J, Rogner M. (1996) Supramolecular architecture of cyanobacterial thylakoid membranes: how is the phycobilisome connected with the photosystems? Photosynth Res 49: 103–118 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
