

SIGIRR, a negative regulator of TLR/IL-1R signalling promotes Microbiota dependent resistance to colonization by enteric bacterial pathogens
- PMID: 23950714
- PMCID: PMC3738496
- DOI: 10.1371/journal.ppat.1003539
SIGIRR, a negative regulator of TLR/IL-1R signalling promotes Microbiota dependent resistance to colonization by enteric bacterial pathogens
Abstract
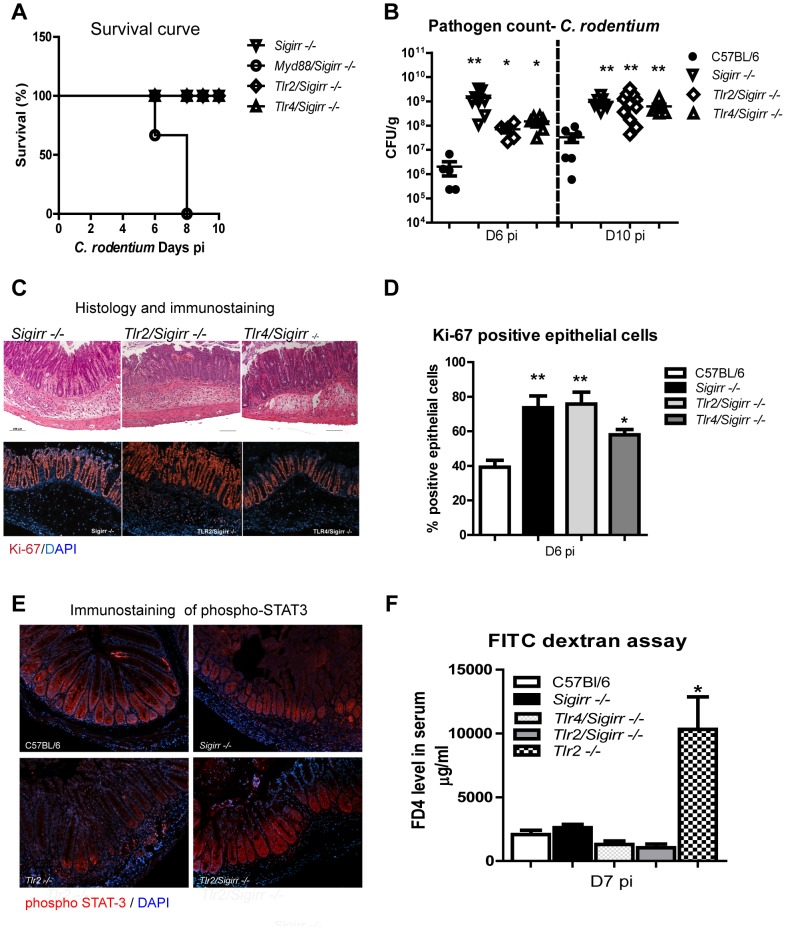
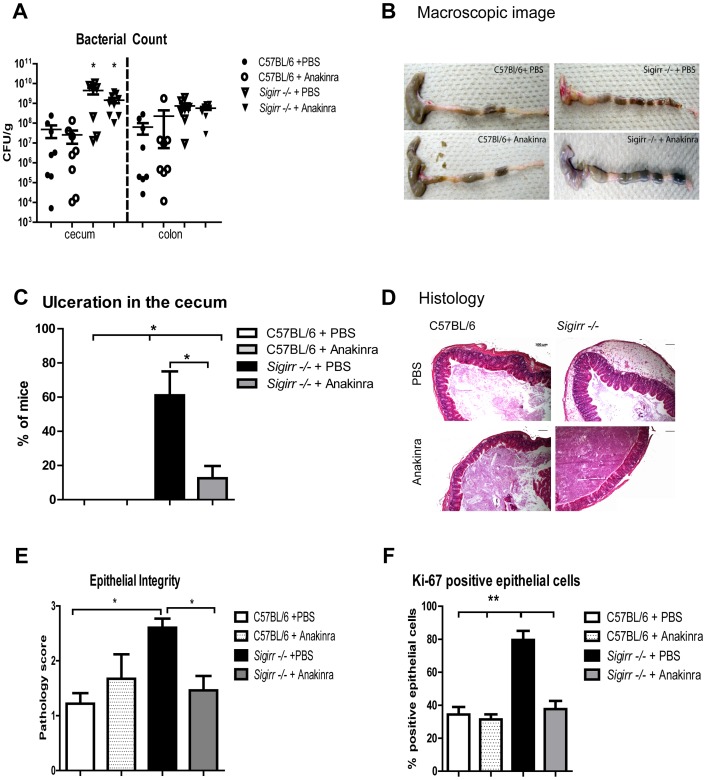
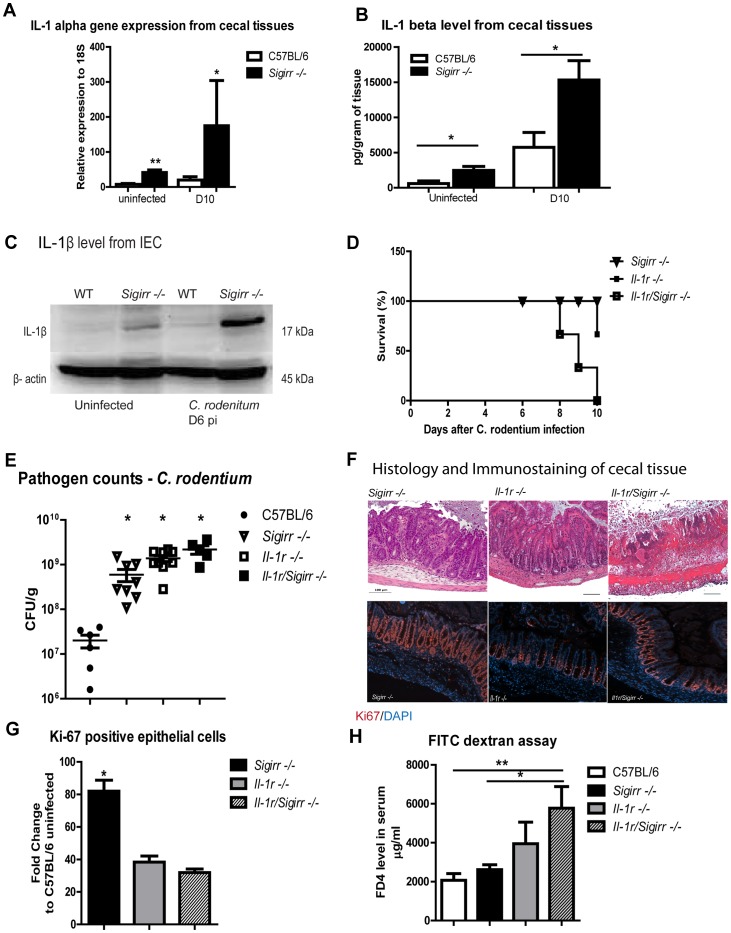
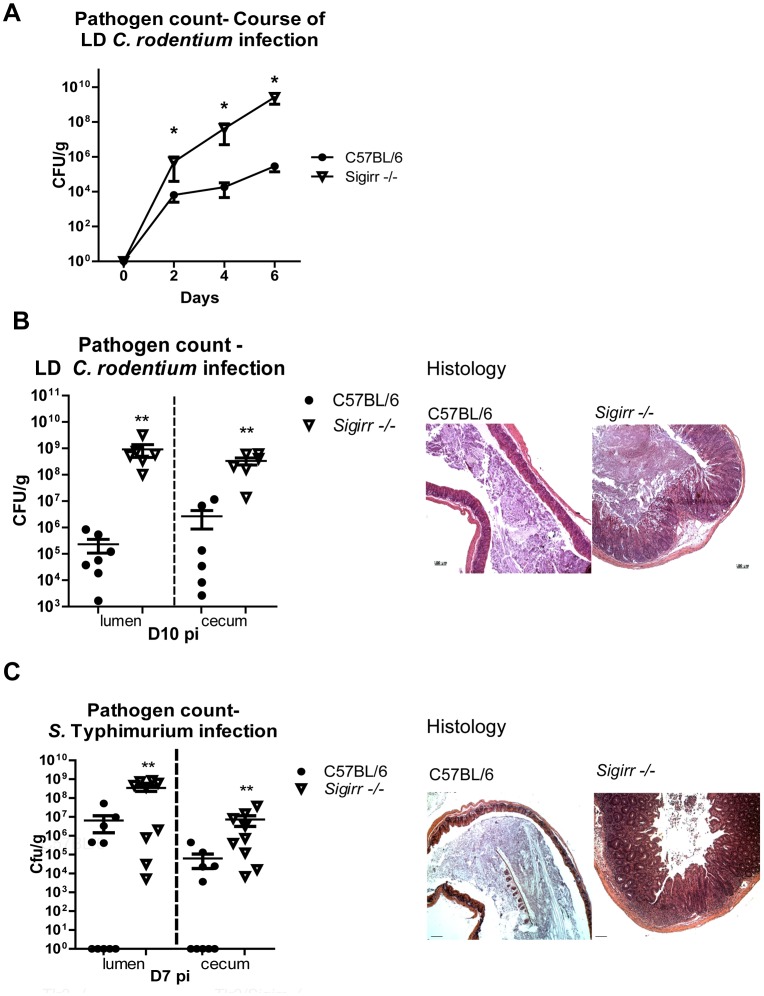
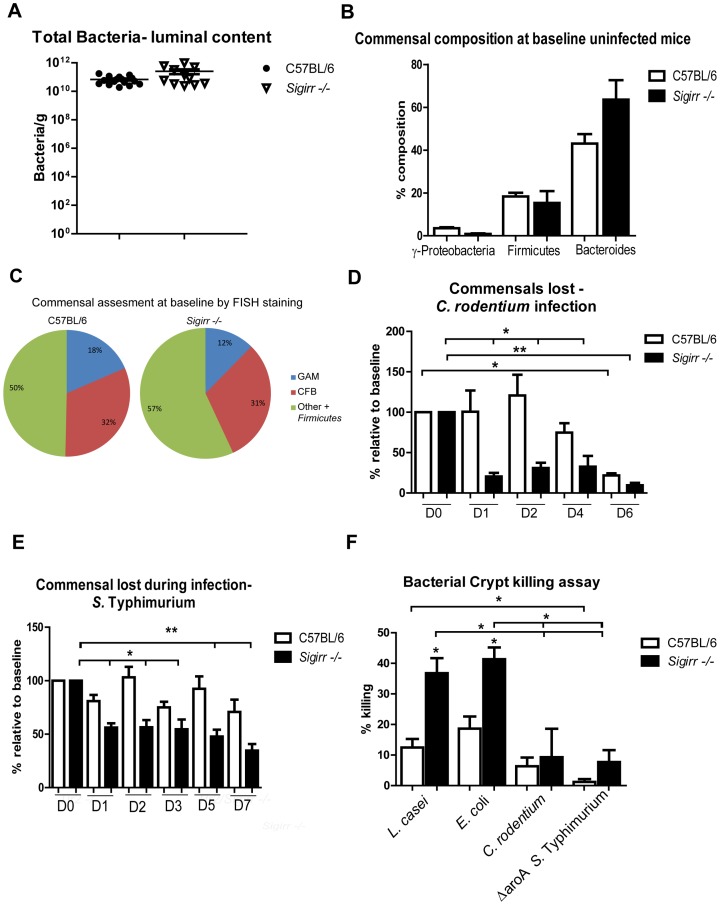
Enteric bacterial pathogens such as enterohemorrhagic E. coli (EHEC) and Salmonella Typhimurium target the intestinal epithelial cells (IEC) lining the mammalian gastrointestinal tract. Despite expressing innate Toll-like receptors (TLRs), IEC are innately hypo-responsive to most bacterial products. This is thought to prevent maladaptive inflammatory responses against commensal bacteria, but it also limits antimicrobial responses by IEC to invading bacterial pathogens, potentially increasing host susceptibility to infection. One reason for the innate hypo-responsiveness of IEC is their expression of Single Ig IL-1 Related Receptor (SIGIRR), a negative regulator of interleukin (IL)-1 and TLR signaling. To address whether SIGIRR expression and the innate hypo-responsiveness of IEC impacts on enteric host defense, Sigirr deficient (-/-) mice were infected with the EHEC related pathogen Citrobacter rodentium. Sigirr -/- mice responded with accelerated IEC proliferation and strong pro-inflammatory and antimicrobial responses but surprisingly, Sigirr -/- mice proved dramatically more susceptible to infection than wildtype mice. Through haematopoietic transplantation studies, it was determined that SIGIRR expression by non-haematopoietic cells (putative IEC) regulated these responses. Moreover, the exaggerated responses were found to be primarily dependent on IL-1R signaling. Whilst exploring the basis for their susceptibility, Sigirr -/- mice were found to be unusually susceptible to intestinal Salmonella Typhimurium colonization, developing enterocolitis without the typical requirement for antibiotic based removal of competing commensal microbes. Strikingly, the exaggerated antimicrobial responses seen in Sigirr -/- mice were found to cause a rapid and dramatic loss of commensal microbes from the infected intestine. This depletion appears to reduce the ability of the microbiota to compete for space and nutrients (colonization resistance) with the invading pathogens, leaving the intestine highly susceptible to pathogen colonization. Thus, SIGIRR expression by IEC reflects a strategy that sacrifices maximal innate responsiveness by IEC in order to promote commensal microbe based colonization resistance against bacterial pathogens.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Mundy R, MacDonald TT, Dougan G, Frankel G, Wiles S (2005) Citrobacter rodentium of mice and man. Cell Microbiol 7: 1697–1706. - PubMed
-
- Bergstrom KS, Sham HP, Zarepour M, Vallance BA (2012) Innate host responses to enteric bacterial pathogens: A balancing act between resistance and tolerance. Cell Microbiol 14(4): 475–84. - PubMed
-
- Lupp C, Robertson ML, Wickham ME, Sekirov I, Champion OL, et al. (2007) Host-mediated inflammation disrupts the intestinal microbiota and promotes the overgrowth of Enterobacteriaceae. Cell Host Microbe 2: 119–129. - PubMed
-
- Gibson DL, Ma C, Bergstrom KS, Huang JT, Man C, et al. (2008) MyD88 signalling plays a critical role in host defence by controlling pathogen burden and promoting epithelial cell homeostasis during Citrobacter rodentium-induced colitis. Cell Microbiol 10: 618–631. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
