

Global analysis of fission yeast mating genes reveals new autophagy factors
- PMID: 23950735
- PMCID: PMC3738441
- DOI: 10.1371/journal.pgen.1003715
Global analysis of fission yeast mating genes reveals new autophagy factors
Abstract
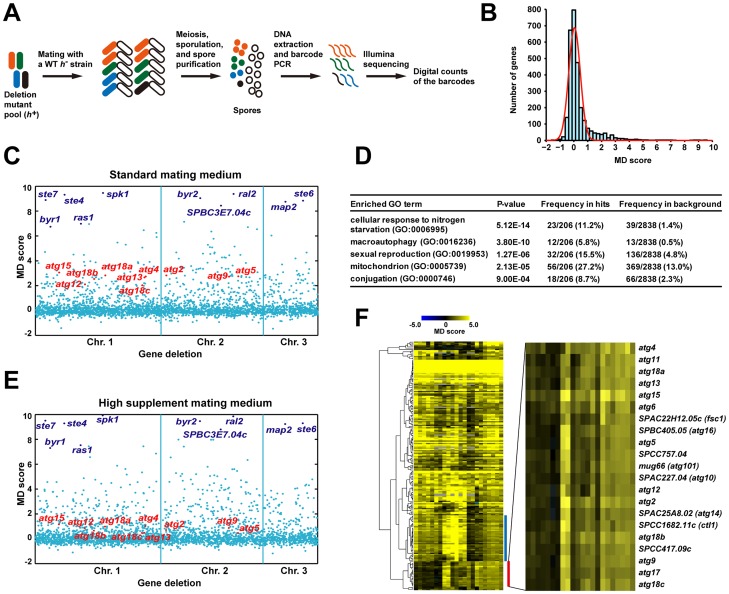
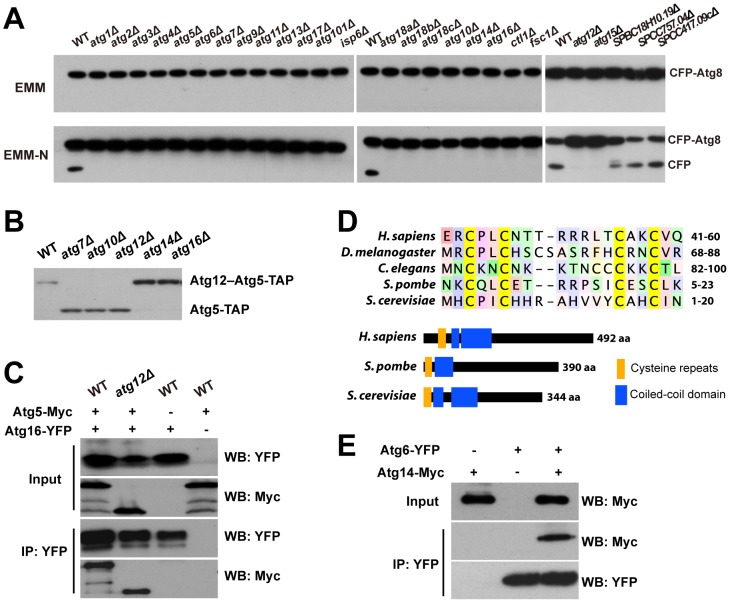
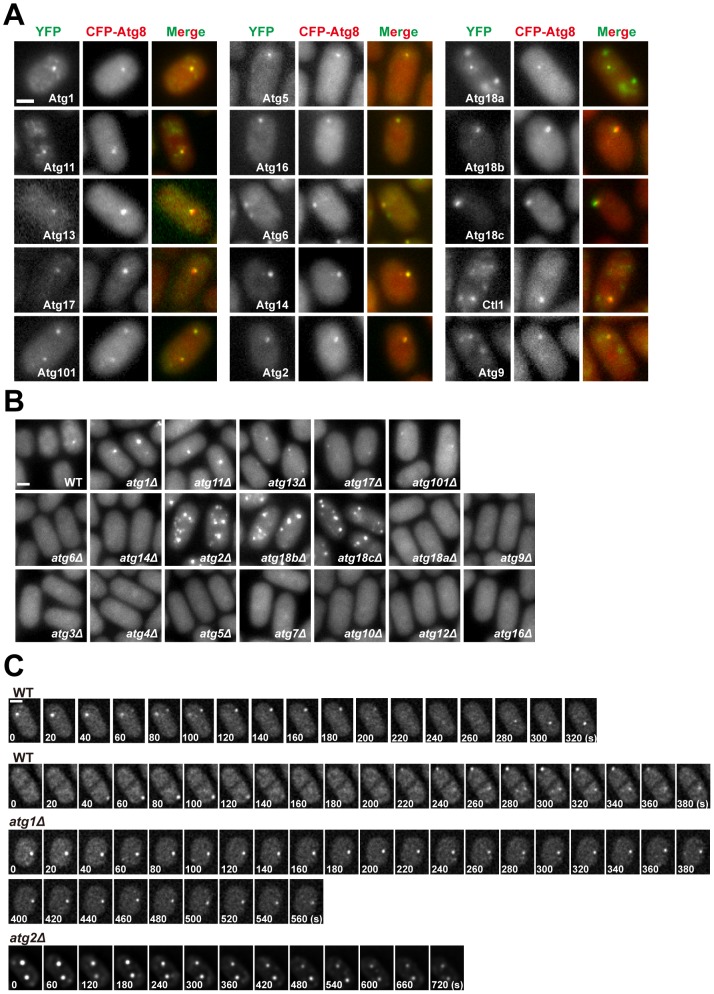
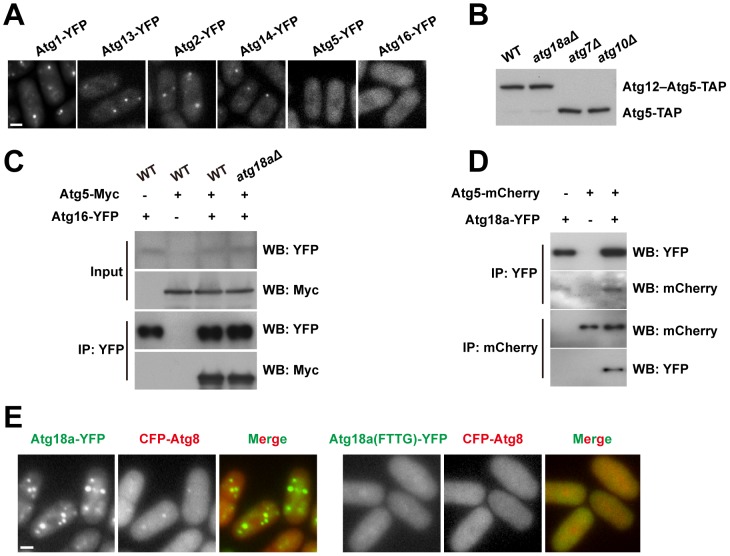
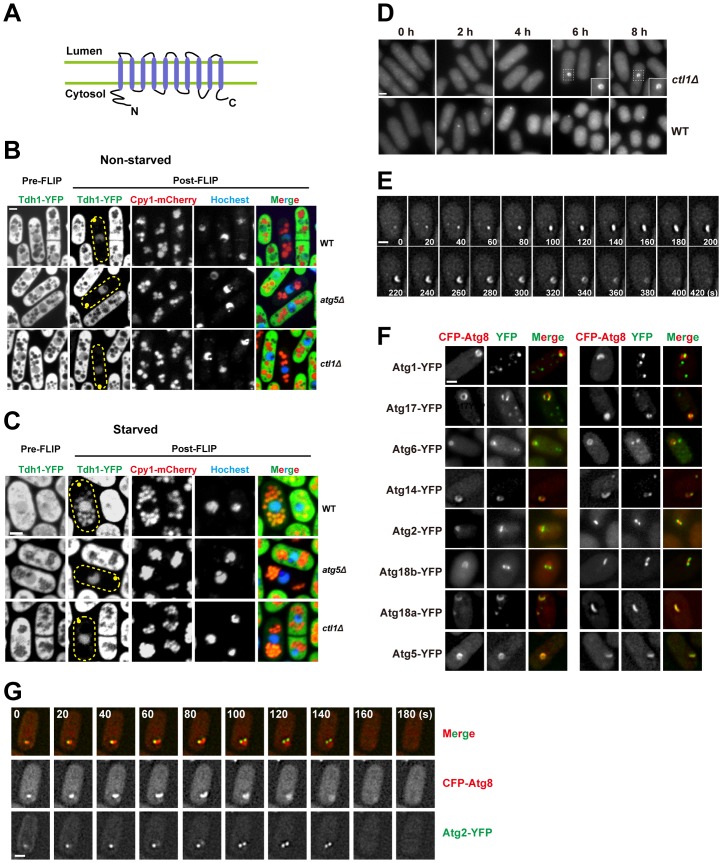
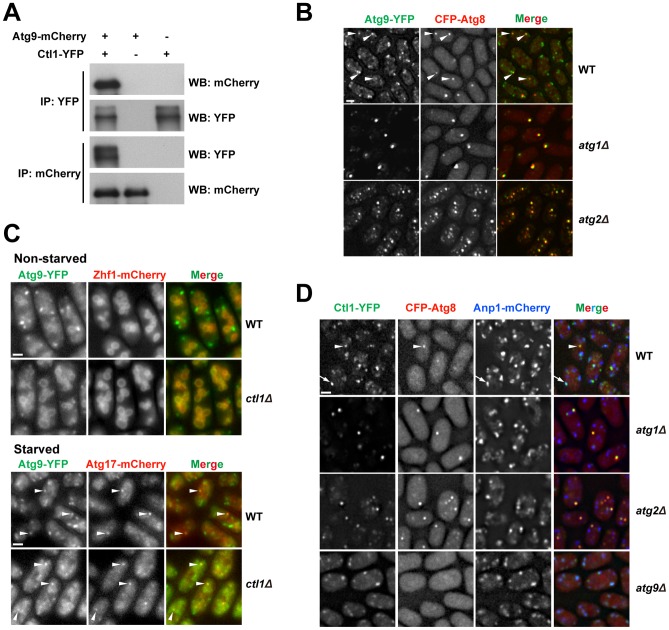
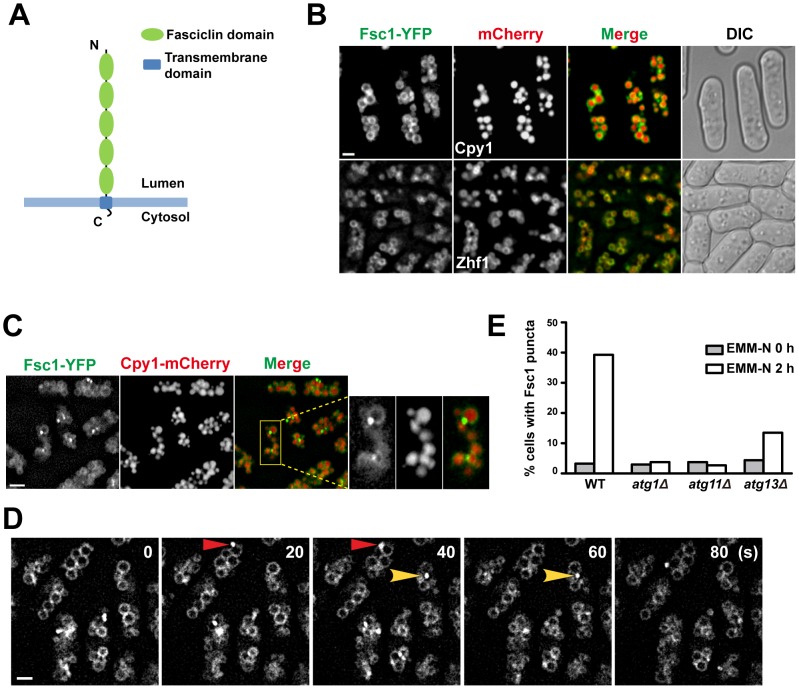
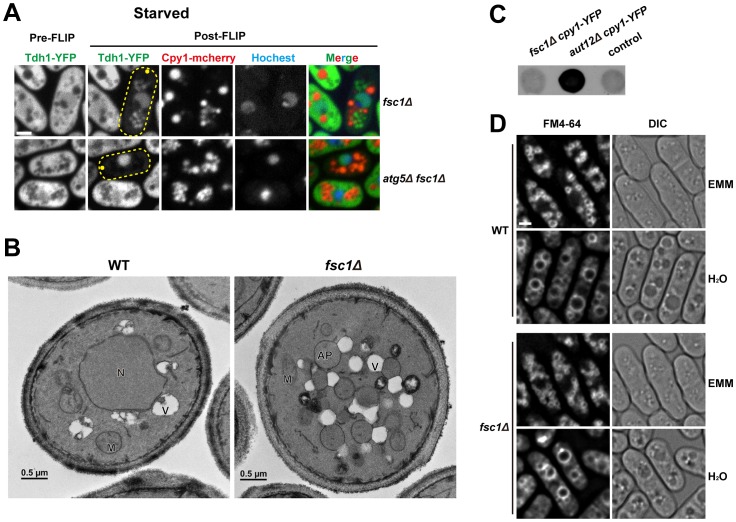
Macroautophagy (autophagy) is crucial for cell survival during starvation and plays important roles in animal development and human diseases. Molecular understanding of autophagy has mainly come from the budding yeast Saccharomyces cerevisiae, and it remains unclear to what extent the mechanisms are the same in other organisms. Here, through screening the mating phenotype of a genome-wide deletion collection of the fission yeast Schizosaccharomyces pombe, we obtained a comprehensive catalog of autophagy genes in this highly tractable organism, including genes encoding three heretofore unidentified core Atg proteins, Atg10, Atg14, and Atg16, and two novel factors, Ctl1 and Fsc1. We systematically examined the subcellular localization of fission yeast autophagy factors for the first time and characterized the phenotypes of their mutants, thereby uncovering both similarities and differences between the two yeasts. Unlike budding yeast, all three Atg18/WIPI proteins in fission yeast are essential for autophagy, and we found that they play different roles, with Atg18a uniquely required for the targeting of the Atg12-Atg5·Atg16 complex. Our investigation of the two novel factors revealed unforeseen autophagy mechanisms. The choline transporter-like protein Ctl1 interacts with Atg9 and is required for autophagosome formation. The fasciclin domain protein Fsc1 localizes to the vacuole membrane and is required for autophagosome-vacuole fusion but not other vacuolar fusion events. Our study sheds new light on the evolutionary diversity of the autophagy machinery and establishes the fission yeast as a useful model for dissecting the mechanisms of autophagy.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Tsukada M, Ohsumi Y (1993) Isolation and characterization of autophagy-defective mutants of Saccharomyces cerevisiae. FEBS Lett 333: 169–174. - PubMed
-
- Kohda TA, Tanaka K, Konomi M, Sato M, Osumi M, et al. (2007) Fission yeast autophagy induced by nitrogen starvation generates a nitrogen source that drives adaptation processes. Genes Cells 12: 155–170 doi:10.1111/j.1365-2443.2007.01041.x - DOI - PubMed
-
- Levine B, Kroemer G (2008) Autophagy in the pathogenesis of disease. Cell 132: 27–42 doi:10.1016/j.cell.2007.12.018 - DOI - PMC - PubMed
-
- Mizushima N, Komatsu M (2011) Autophagy: renovation of cells and tissues. Cell 147: 728–741 doi:10.1016/j.cell.2011.10.026 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
