

MicroRNA-185 and 342 inhibit tumorigenicity and induce apoptosis through blockade of the SREBP metabolic pathway in prostate cancer cells
- PMID: 23951060
- PMCID: PMC3739799
- DOI: 10.1371/journal.pone.0070987
MicroRNA-185 and 342 inhibit tumorigenicity and induce apoptosis through blockade of the SREBP metabolic pathway in prostate cancer cells
Abstract
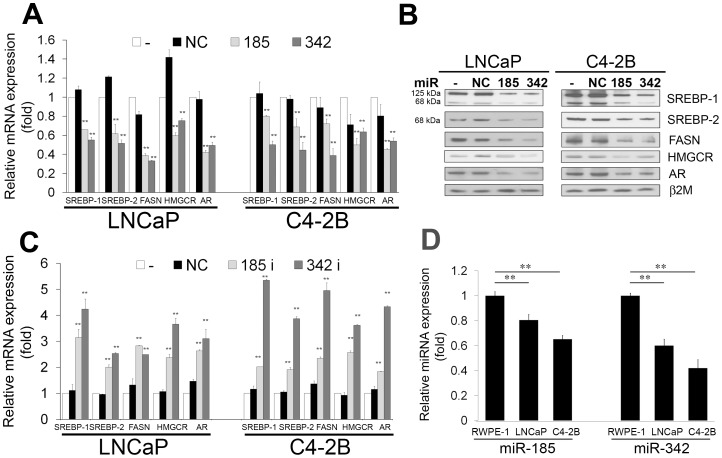
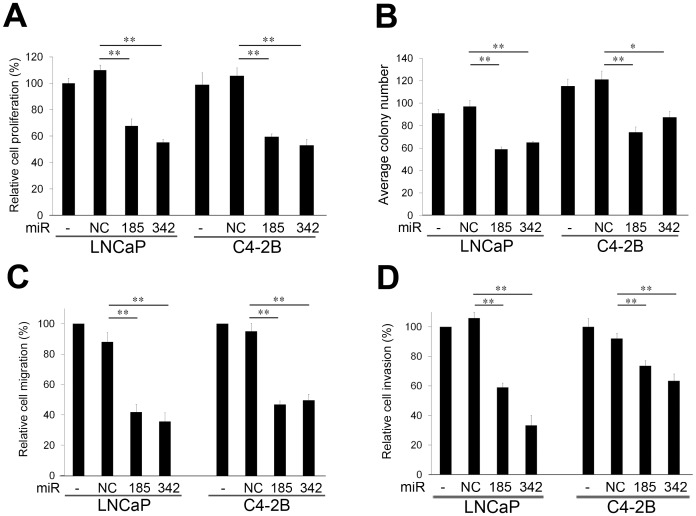
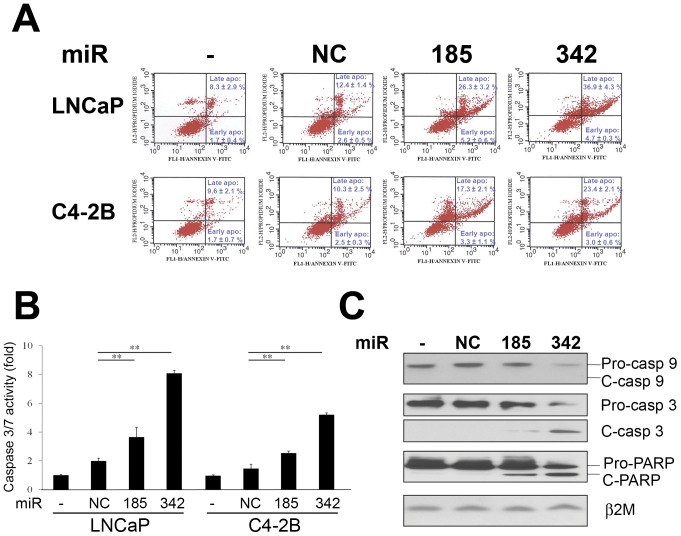
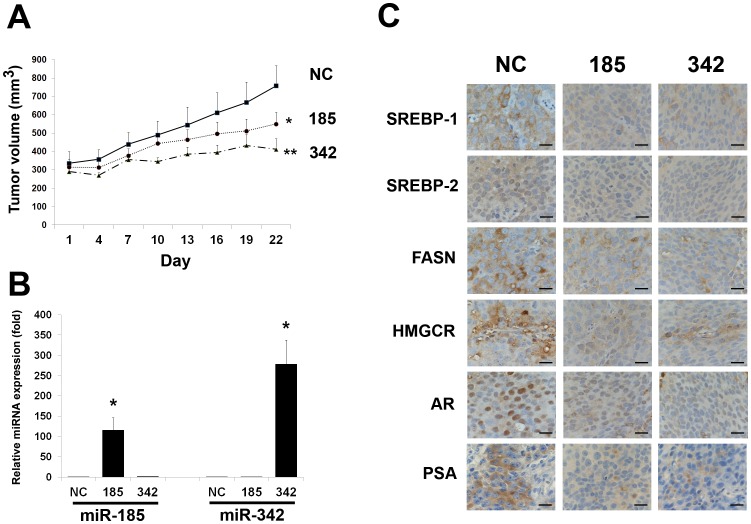
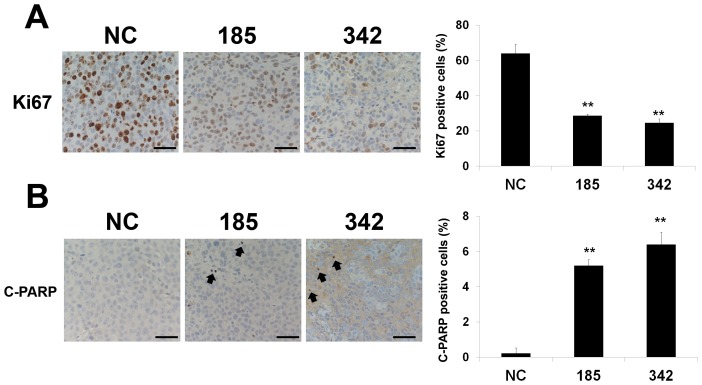
MicroRNA (miRNA or miR) inhibition of oncogenic related pathways has been shown to be a promising therapeutic approach for cancer. Aberrant lipid and cholesterol metabolism is involved in prostate cancer development and progression to end-stage disease. We recently demonstrated that a key transcription factor for lipogenesis, sterol regulatory element-binding protein-1 (SREBP-1), induced fatty acid and lipid accumulation and androgen receptor (AR) transcriptional activity, and also promoted prostate cancer cell growth and castration resistance. SREBP-1 was overexpressed in human prostate cancer and castration-resistant patient specimens. These experimental and clinical results indicate that SREBP-1 is a potential oncogenic transcription factor in prostate cancer. In this study, we identified two miRNAs, miR-185 and 342, that control lipogenesis and cholesterogenesis in prostate cancer cells by inhibiting SREBP-1 and 2 expression and down-regulating their targeted genes, including fatty acid synthase (FASN) and 3-hydroxy-3-methylglutaryl CoA reductase (HMGCR). Both miR-185 and 342 inhibited tumorigenicity, cell growth, migration and invasion in prostate cancer cell culture and xenograft models coincident with their blockade of lipogenesis and cholesterogenesis. Intrinsic miR-185 and 342 expression was significantly decreased in prostate cancer cells compared to non-cancerous epithelial cells. Restoration of miR-185 and 342 led to caspase-dependent apoptotic death in prostate cancer cells. The newly identified miRNAs, miR-185 and 342, represent a novel targeting mechanism for prostate cancer therapy.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
