

Weight-bearing locomotion in the developing opossum, Monodelphis domestica following spinal transection: remodeling of neuronal circuits caudal to lesion
- PMID: 23951105
- PMCID: PMC3741377
- DOI: 10.1371/journal.pone.0071181
Weight-bearing locomotion in the developing opossum, Monodelphis domestica following spinal transection: remodeling of neuronal circuits caudal to lesion
Abstract
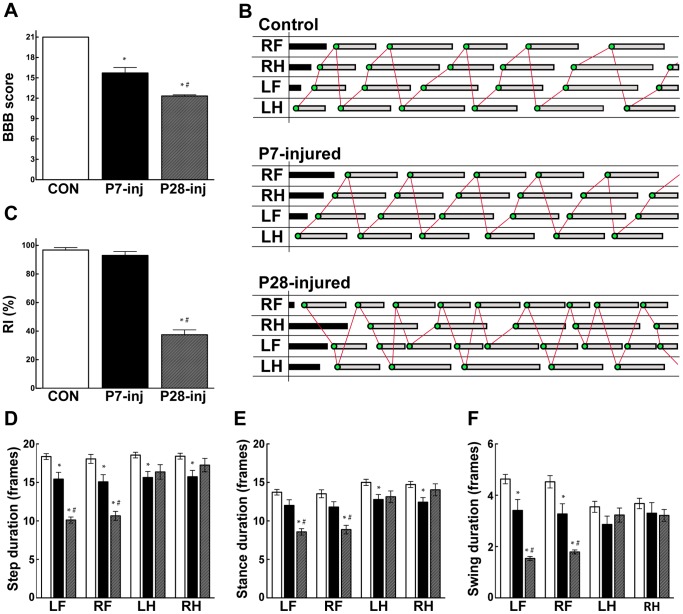
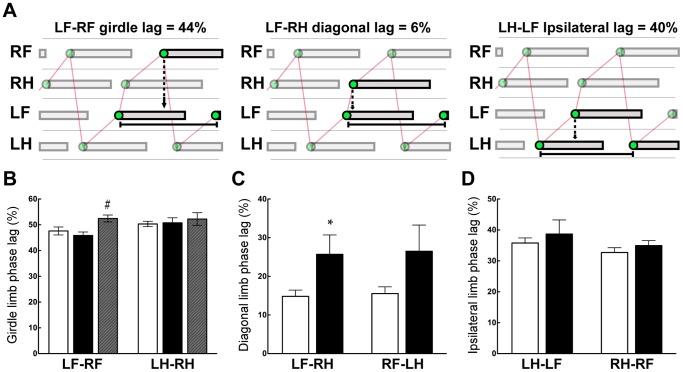
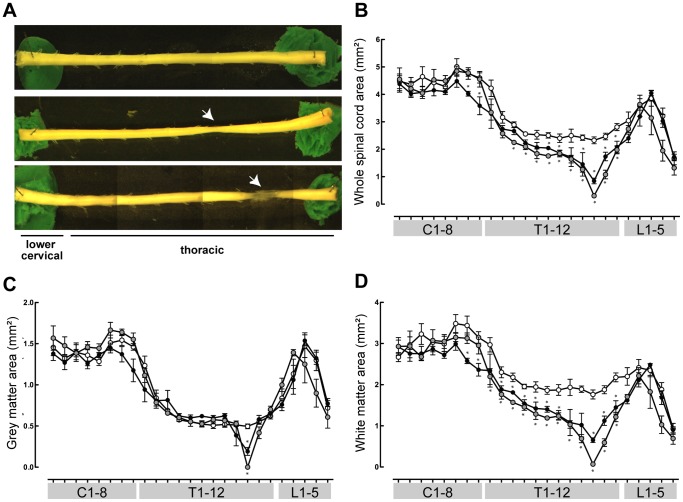
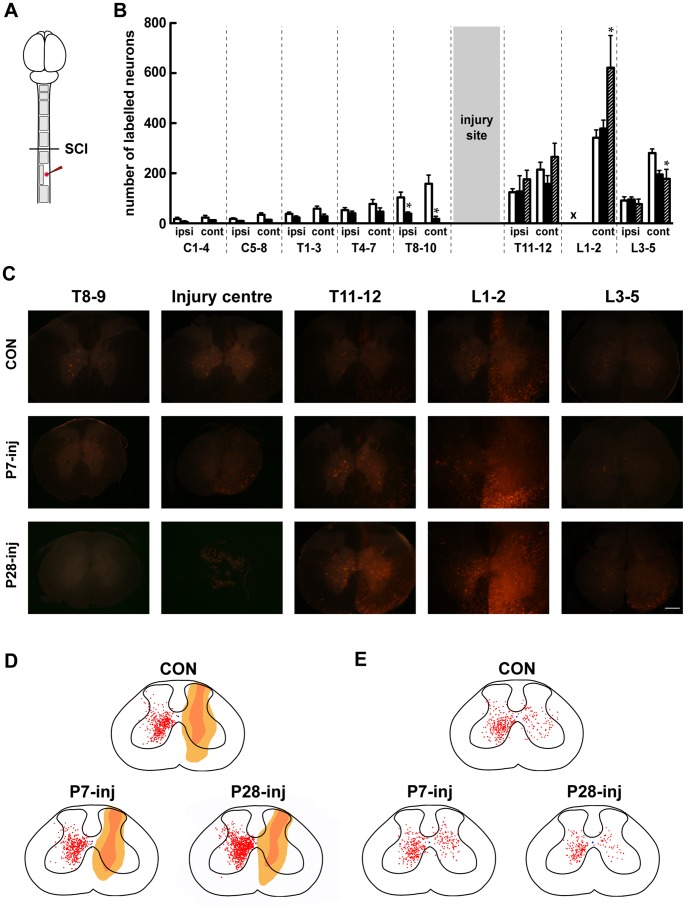
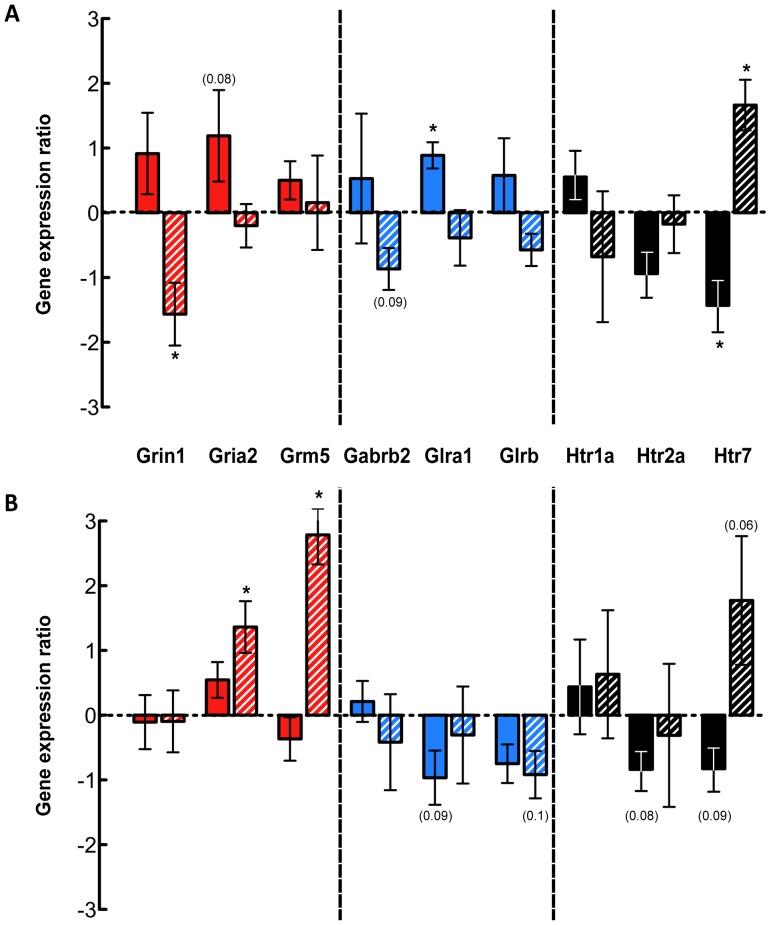
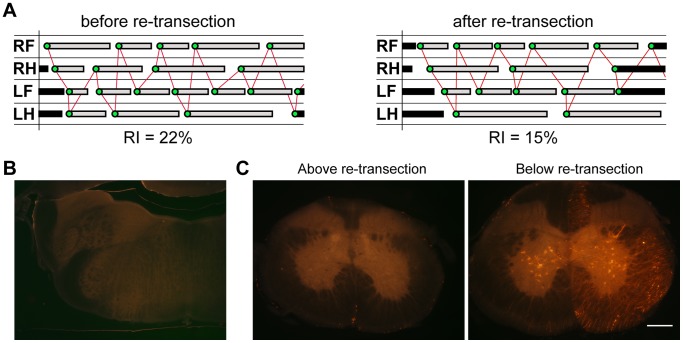
Complete spinal transection in the mature nervous system is typically followed by minimal axonal repair, extensive motor paralysis and loss of sensory functions caudal to the injury. In contrast, the immature nervous system has greater capacity for repair, a phenomenon sometimes called the infant lesion effect. This study investigates spinal injuries early in development using the marsupial opossum Monodelphis domestica whose young are born very immature, allowing access to developmental stages only accessible in utero in eutherian mammals. Spinal cords of Monodelphis pups were completely transected in the lower thoracic region, T10, on postnatal-day (P)7 or P28 and the animals grew to adulthood. In P7-injured animals regrown supraspinal and propriospinal axons through the injury site were demonstrated using retrograde axonal labelling. These animals recovered near-normal coordinated overground locomotion, but with altered gait characteristics including foot placement phase lags. In P28-injured animals no axonal regrowth through the injury site could be demonstrated yet they were able to perform weight-supporting hindlimb stepping overground and on the treadmill. When placed in an environment of reduced sensory feedback (swimming) P7-injured animals swam using their hindlimbs, suggesting that the axons that grew across the lesion made functional connections; P28-injured animals swam using their forelimbs only, suggesting that their overground hindlimb movements were reflex-dependent and thus likely to be generated locally in the lumbar spinal cord. Modifications to propriospinal circuitry in P7- and P28-injured opossums were demonstrated by changes in the number of fluorescently labelled neurons detected in the lumbar cord following tracer studies and changes in the balance of excitatory, inhibitory and neuromodulatory neurotransmitter receptors' gene expression shown by qRT-PCR. These results are discussed in the context of studies indicating that although following injury the isolated segment of the spinal cord retains some capability of rhythmic movement the mechanisms involved in weight-bearing locomotion are distinct.
Conflict of interest statement
Figures
Similar articles
-
Age-dependent transcriptome and proteome following transection of neonatal spinal cord of Monodelphis domestica (South American grey short-tailed opossum).PLoS One. 2014 Jun 10;9(6):e99080. doi: 10.1371/journal.pone.0099080. eCollection 2014. PLoS One. 2014. PMID: 24914927 Free PMC article.
-
Spontaneous development of full weight-supported stepping after complete spinal cord transection in the neonatal opossum, Monodelphis domestica.PLoS One. 2011;6(11):e26826. doi: 10.1371/journal.pone.0026826. Epub 2011 Nov 2. PLoS One. 2011. PMID: 22073202 Free PMC article.
-
Transplants and neurotrophic factors increase regeneration and recovery of function after spinal cord injury.Prog Brain Res. 2002;137:257-73. doi: 10.1016/s0079-6123(02)37020-1. Prog Brain Res. 2002. PMID: 12440372 Review.
-
Age-related differences in the local cellular and molecular responses to injury in developing spinal cord of the opossum, Monodelphis domestica.Eur J Neurosci. 2007 Mar;25(6):1725-42. doi: 10.1111/j.1460-9568.2007.05439.x. Eur J Neurosci. 2007. PMID: 17432961
-
Spinal repair in immature animals: a novel approach using the South American opossum Monodelphis domestica.Clin Exp Pharmacol Physiol. 2000 Jul;27(7):542-7. doi: 10.1046/j.1440-1681.2000.03296.x. Clin Exp Pharmacol Physiol. 2000. PMID: 10874515 Review.
Cited by
-
Age-dependent transcriptome and proteome following transection of neonatal spinal cord of Monodelphis domestica (South American grey short-tailed opossum).PLoS One. 2014 Jun 10;9(6):e99080. doi: 10.1371/journal.pone.0099080. eCollection 2014. PLoS One. 2014. PMID: 24914927 Free PMC article.
-
A bipedal mammalian model for spinal cord injury research: The tammar wallaby.F1000Res. 2017 Jun 15;6:921. doi: 10.12688/f1000research.11712.1. eCollection 2017. F1000Res. 2017. PMID: 28721206 Free PMC article.
-
Comparison of 6 Injectable Anesthetic Regimens and Isoflurane in Gray Short-tailed Opossums (Monodelphis domestica).J Am Assoc Lab Anim Sci. 2017 Sep 1;56(5):544-549. J Am Assoc Lab Anim Sci. 2017. PMID: 28903826 Free PMC article. Clinical Trial.
-
ABC Efflux Transporters and Solute Carriers in the Early Developing Brain of a Marsupial Monodelphis domestica (South American Gray Short-Tailed Opossum).J Comp Neurol. 2024 Jul;532(7):e25655. doi: 10.1002/cne.25655. J Comp Neurol. 2024. PMID: 38980080 Free PMC article.
-
Treadmill exercise with bone marrow stromal cells transplantation facilitates neuroprotective effect through BDNF-ERK1/2 pathway in spinal cord injury rats.J Exerc Rehabil. 2018 Jun 30;14(3):335-340. doi: 10.12965/jer.1836264.132. eCollection 2018 Jun. J Exerc Rehabil. 2018. PMID: 30018915 Free PMC article.
References
-
- Steward O, Popovich PG, Dietrich WD, Kleitman N (2012) Replication and reproducibility in spinal cord injury research. Experimental neurology 233: 597–605. - PubMed
-
- Sherrington CS (1906) The integrative action of the nervous system: New Haven: Yale University Press.
-
- Brown TG (1911) The intrinsic factors in the act of progression in the mammal. Proc R Soc Lond B Biol Sci 84: 308–319.
-
- Barbeau H, Rossignol S (1987) Recovery of locomotion after chronic spinalization in the adult cat. Brain research 412: 84–95. - PubMed
-
- Lovely RG, Gregor RJ, Roy RR, Edgerton VR (1990) Weight-bearing hindlimb stepping in treadmill-exercised adult spinal cats. Brain research 514: 206–218. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
