

Histone deacetylase SIRT1 modulates and deacetylates DNA base excision repair enzyme thymine DNA glycosylase
- PMID: 23952905
- PMCID: PMC4465237
- DOI: 10.1042/BJ20130670
Histone deacetylase SIRT1 modulates and deacetylates DNA base excision repair enzyme thymine DNA glycosylase
Abstract
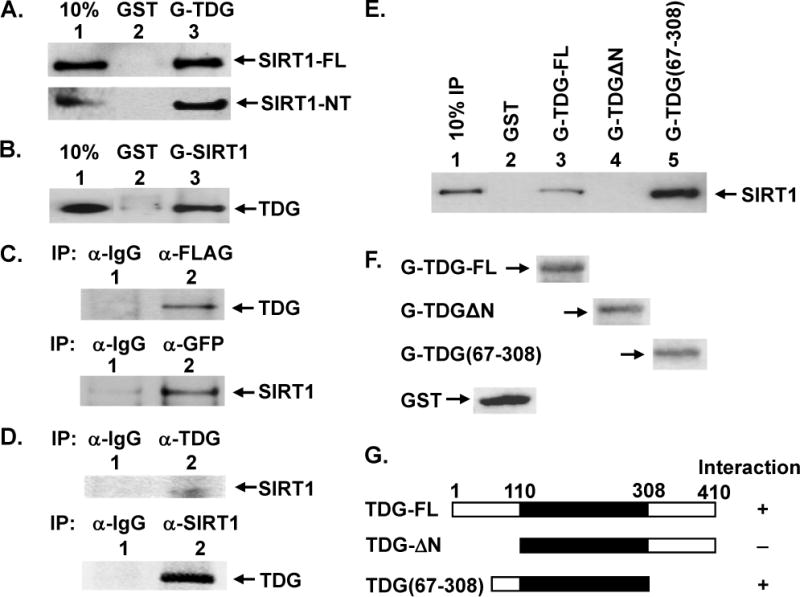
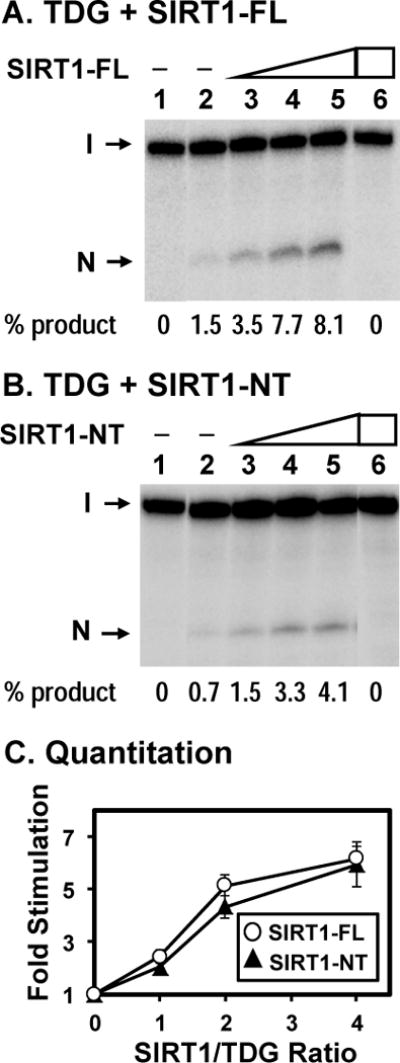
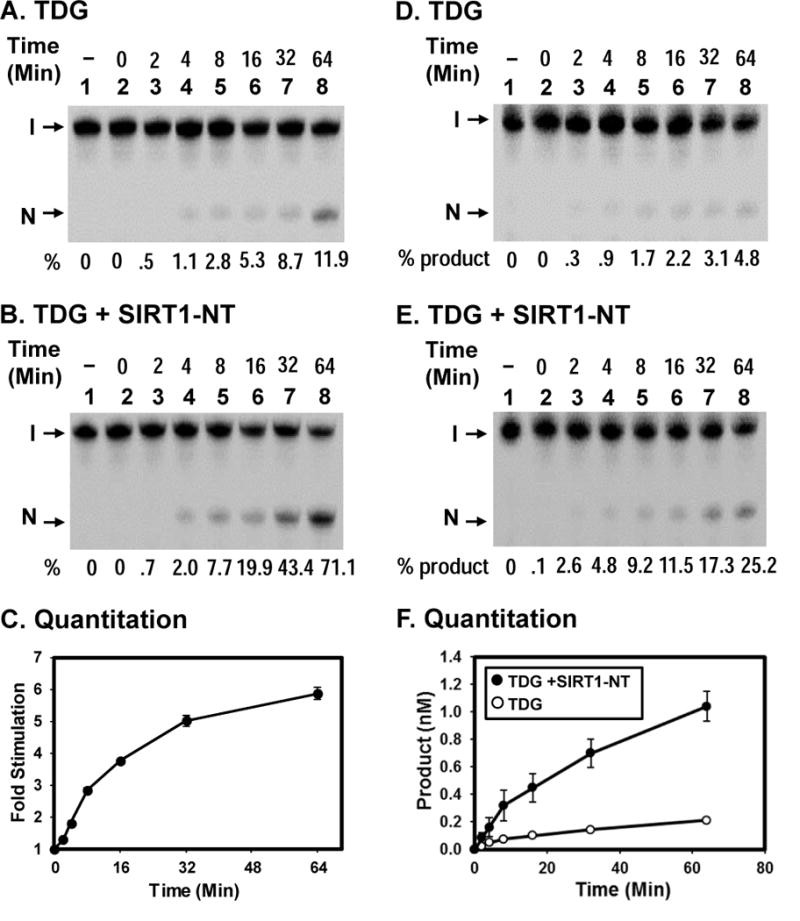
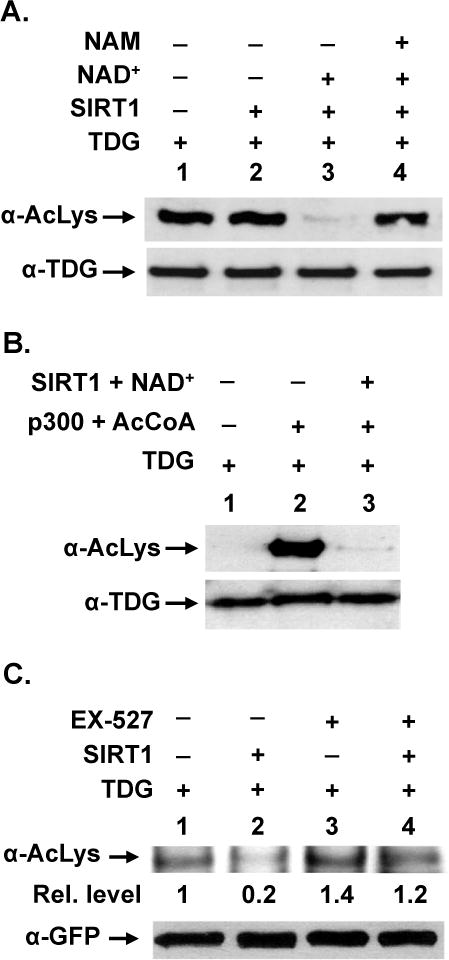
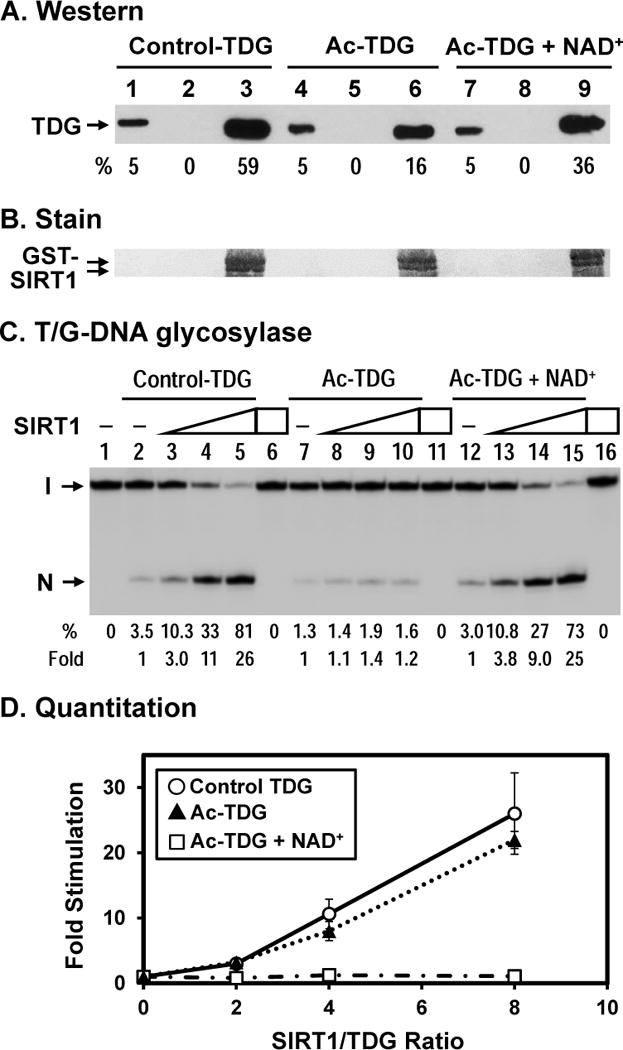
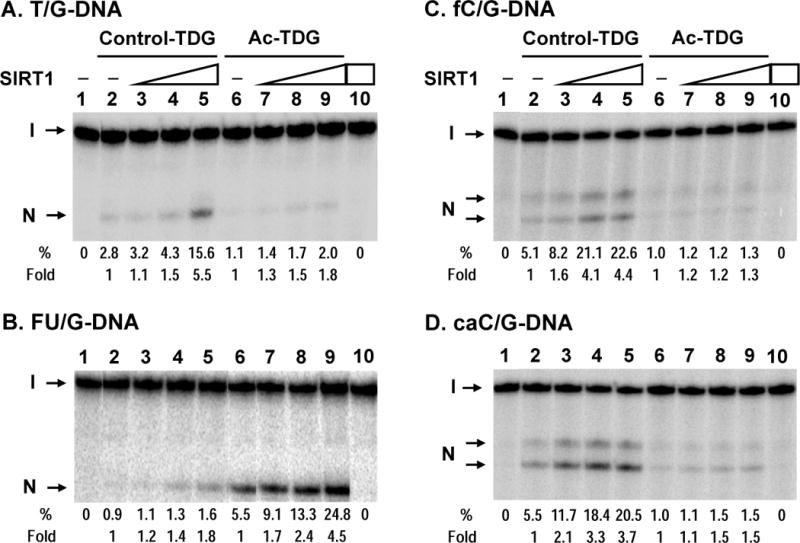
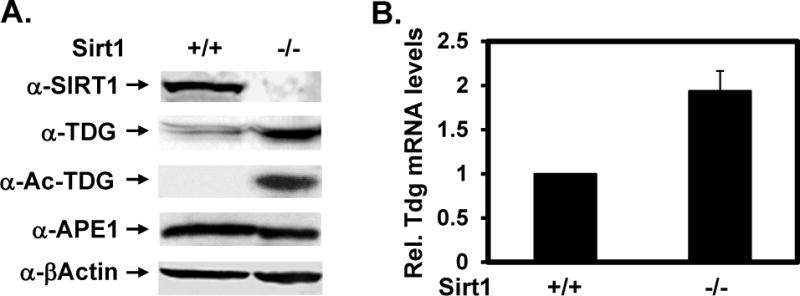
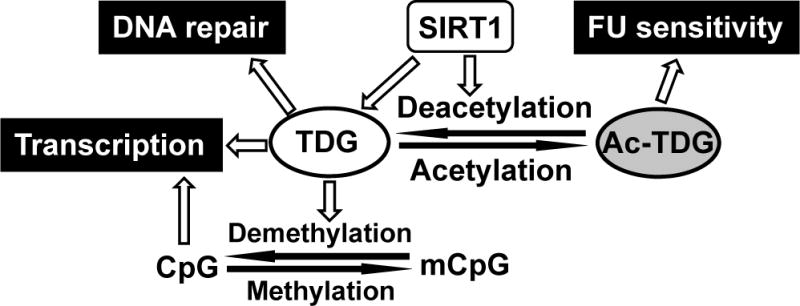
TDG (thymine DNA glycosylase) is an essential multifunctional enzyme involved in DNA base excision repair, DNA demethylation and transcription regulation. TDG is the predominant enzyme that removes thymine from T/G mispair, which arises due to deamination of 5-methyl-cytosine at the CpG dinucleotide, thereby preventing C to T mutations. SIRT1 is a member of class III NAD+-dependent histone/protein deacetylases. In the present study, we demonstrate that SIRT1 interacts with residues 67-110 of hTDG (human TDG). In addition, SIRT1 enhances TDG glycosylase activity and deacetylates acetylated TDG. TDG acetylation weakens its interaction with SIRT1. Although acetylated TDG has reduced glycosylase activity towards T/G, 5-formylcytosine/G and 5-carboxylcytosine/G, it has a stronger activity towards a 5-fluorouracil/G substrate as compared with unmodified TDG. SIRT1 weakly stimulates acetylated hTDG activity towards T/G, 5-formylcytosine/G and 5-carboxylcytosine/G as compared with control hTDG. Sirt1-knockout mouse embryonic fibroblast cells have higher levels of TDG expression and acetylation. The physical and functional interactions between SIRT1 and TDG may mediate DNA repair, gene expression and FU (5-fluorouracil)-mediated cytotoxicity.
Figures
Similar articles
-
Thymine DNA glycosylase can rapidly excise 5-formylcytosine and 5-carboxylcytosine: potential implications for active demethylation of CpG sites.J Biol Chem. 2011 Oct 14;286(41):35334-35338. doi: 10.1074/jbc.C111.284620. Epub 2011 Aug 23. J Biol Chem. 2011. PMID: 21862836 Free PMC article.
-
Lesion processing by a repair enzyme is severely curtailed by residues needed to prevent aberrant activity on undamaged DNA.Proc Natl Acad Sci U S A. 2012 May 22;109(21):8091-6. doi: 10.1073/pnas.1201010109. Epub 2012 May 9. Proc Natl Acad Sci U S A. 2012. PMID: 22573813 Free PMC article.
-
Sumoylation of thymine DNA glycosylase impairs productive binding to substrate sites in DNA.J Biol Chem. 2024 Nov;300(11):107902. doi: 10.1016/j.jbc.2024.107902. Epub 2024 Oct 18. J Biol Chem. 2024. PMID: 39426728 Free PMC article.
-
The enigmatic thymine DNA glycosylase.DNA Repair (Amst). 2007 Apr 1;6(4):489-504. doi: 10.1016/j.dnarep.2006.10.013. Epub 2006 Nov 20. DNA Repair (Amst). 2007. PMID: 17116428 Review.
-
Role of base excision repair in maintaining the genetic and epigenetic integrity of CpG sites.DNA Repair (Amst). 2015 Aug;32:33-42. doi: 10.1016/j.dnarep.2015.04.011. Epub 2015 May 1. DNA Repair (Amst). 2015. PMID: 26021671 Free PMC article. Review.
Cited by
-
The multifaceted influence of histone deacetylases on DNA damage signalling and DNA repair.Nucleic Acids Res. 2016 Dec 1;44(21):10017-10030. doi: 10.1093/nar/gkw922. Epub 2016 Oct 13. Nucleic Acids Res. 2016. PMID: 27738139 Free PMC article. Review.
-
The dual role of sirtuins in cancer: biological functions and implications.Front Oncol. 2024 Jun 14;14:1384928. doi: 10.3389/fonc.2024.1384928. eCollection 2024. Front Oncol. 2024. PMID: 38947884 Free PMC article. Review.
-
SIRT6 protein deacetylase interacts with MYH DNA glycosylase, APE1 endonuclease, and Rad9-Rad1-Hus1 checkpoint clamp.BMC Mol Biol. 2015 Jun 11;16:12. doi: 10.1186/s12867-015-0041-9. BMC Mol Biol. 2015. PMID: 26063178 Free PMC article.
-
Histone/protein deacetylase SIRT1 is an anticancer therapeutic target.Am J Cancer Res. 2014 May 26;4(3):211-21. eCollection 2014. Am J Cancer Res. 2014. PMID: 24959376 Free PMC article.
-
Defective Base Excision Repair of Oxidative DNA Damage in Vascular Smooth Muscle Cells Promotes Atherosclerosis.Circulation. 2018 Oct 2;138(14):1446-1462. doi: 10.1161/CIRCULATIONAHA.117.033249. Circulation. 2018. PMID: 29643057 Free PMC article.
References
-
- Krokan HE, Nilsen H, Skorpen F, Otterlei M, Slupphaug G. Base excision repair of DNA in mammalian cells. FEBS Lett. 2000;476:73–77. - PubMed
-
- Cortazar D, Kunz C, Saito Y, Steinacher R, Schar P. The enigmatic thymine DNA glycosylase. DNA Repair (Amst) 2007;6:489–504. - PubMed
-
- Hardeland U, Bentele M, Lettieri T, Steinacher R, Jiricny J, Schar P. Thymine DNA glycosylase. Prog Nucleic Acid Res Mol Biol. 2001;68:235–253. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous