

Developmental fate and cellular maturity encoded in human regulatory DNA landscapes
- PMID: 23953118
- PMCID: PMC3962256
- DOI: 10.1016/j.cell.2013.07.020
Developmental fate and cellular maturity encoded in human regulatory DNA landscapes
Abstract
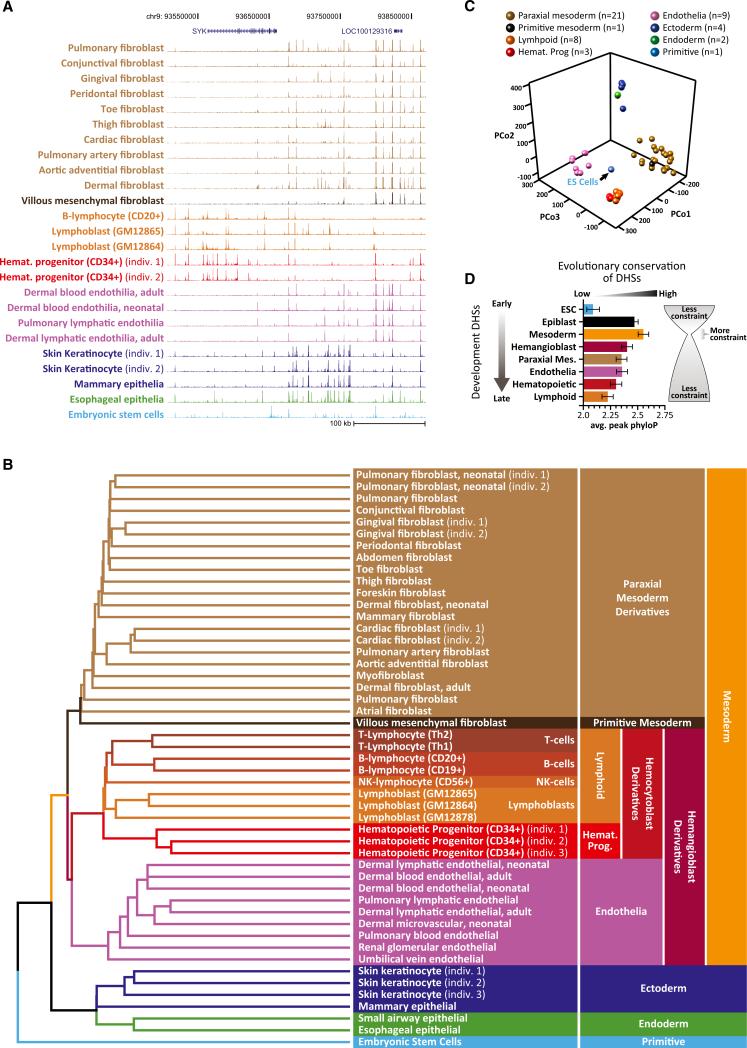
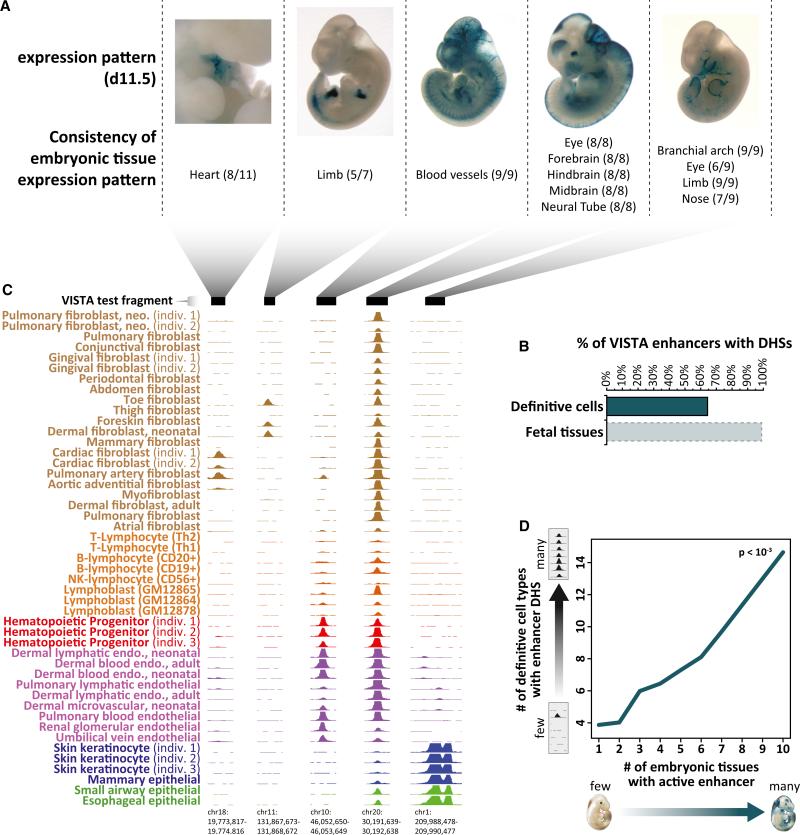
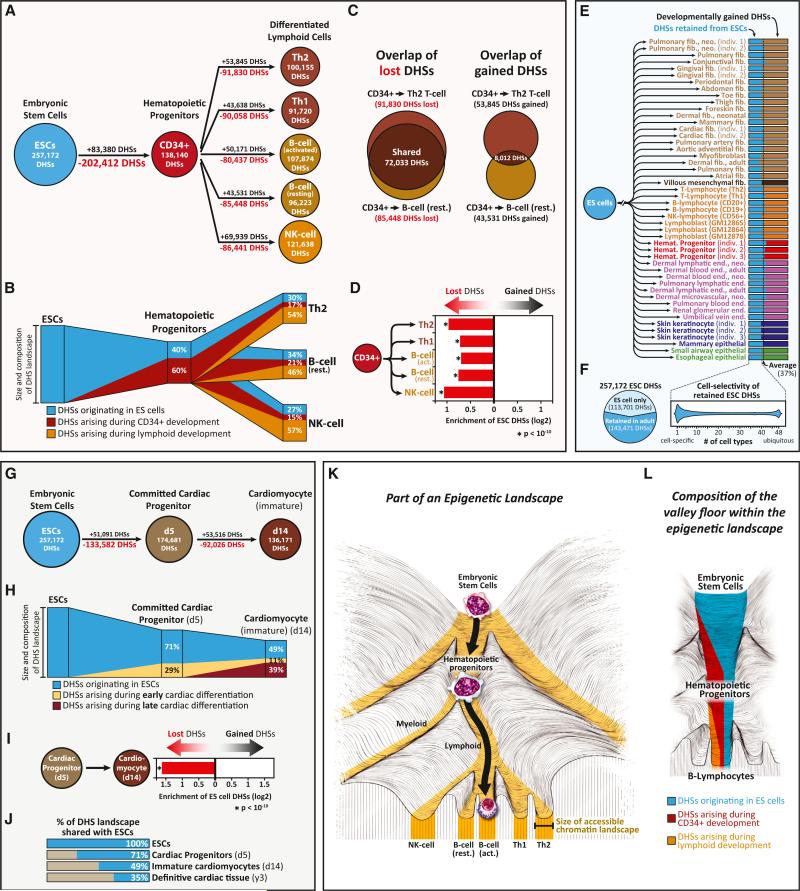
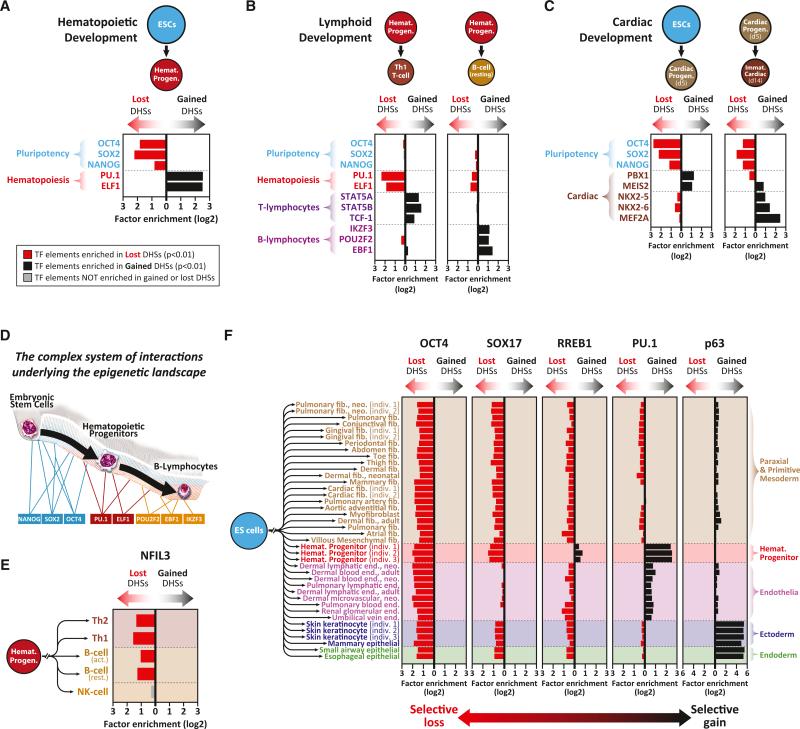
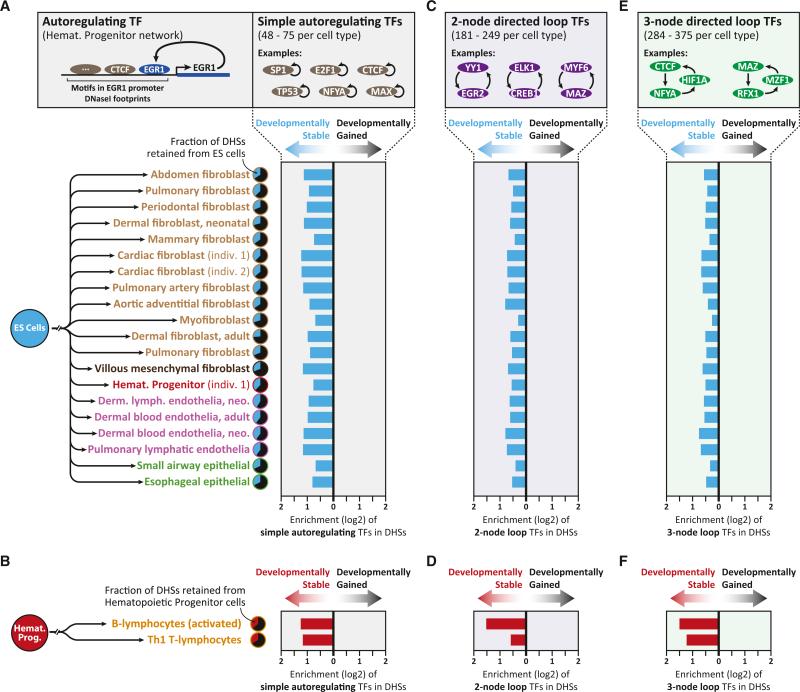
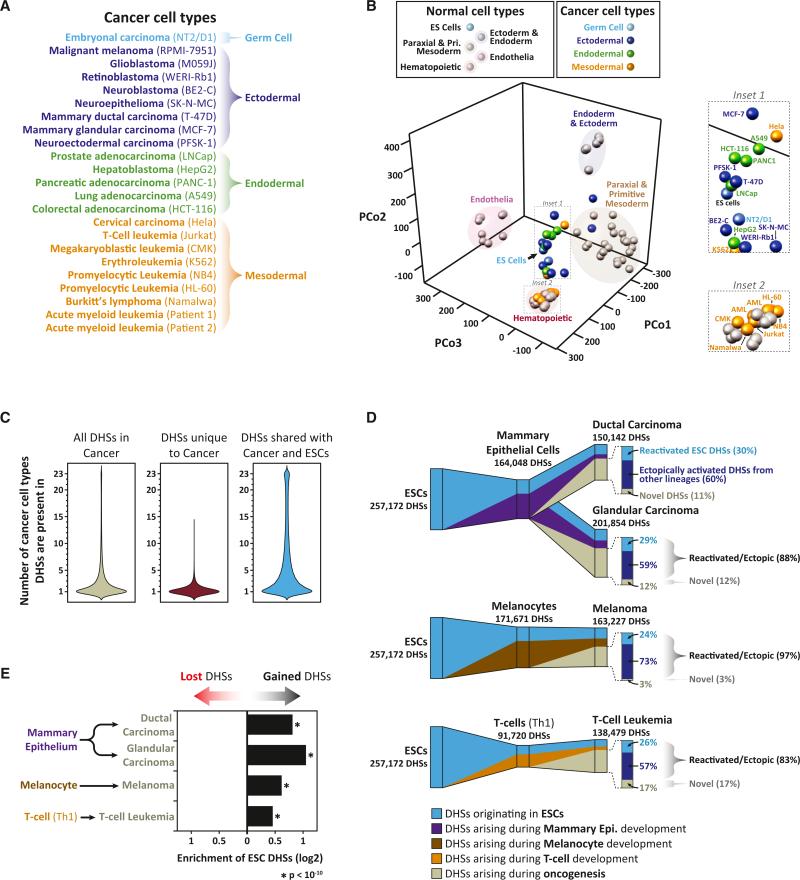
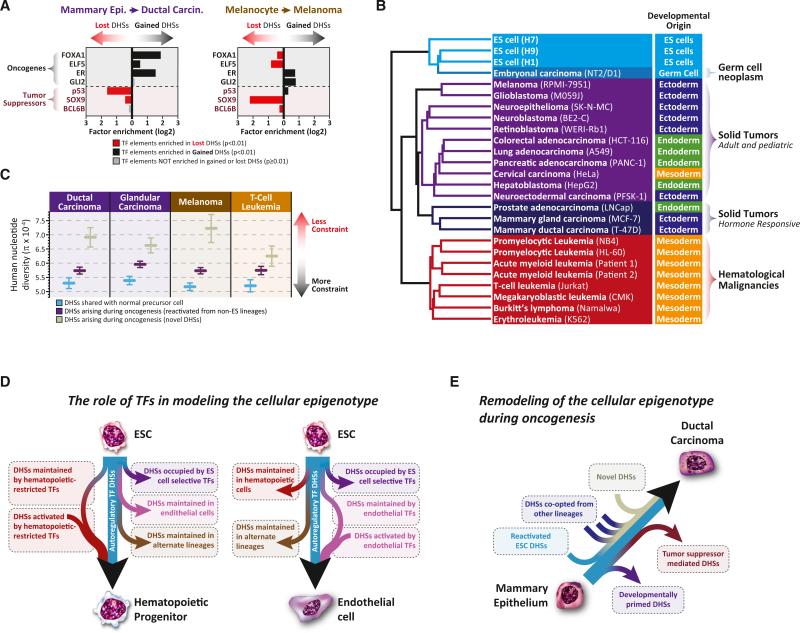
Cellular-state information between generations of developing cells may be propagated via regulatory regions. We report consistent patterns of gain and loss of DNase I-hypersensitive sites (DHSs) as cells progress from embryonic stem cells (ESCs) to terminal fates. DHS patterns alone convey rich information about cell fate and lineage relationships distinct from information conveyed by gene expression. Developing cells share a proportion of their DHS landscapes with ESCs; that proportion decreases continuously in each cell type as differentiation progresses, providing a quantitative benchmark of developmental maturity. Developmentally stable DHSs densely encode binding sites for transcription factors involved in autoregulatory feedback circuits. In contrast to normal cells, cancer cells extensively reactivate silenced ESC DHSs and those from developmental programs external to the cell lineage from which the malignancy derives. Our results point to changes in regulatory DNA landscapes as quantitative indicators of cell-fate transitions, lineage relationships, and dysfunction.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
References
-
- Alon U. An Introduction to Systems Biology: Design Principles of Biological Circuits. First Edition Chapman and Hall/CRC; London: 2006.
-
- Beard J. Embryological aspects and etiology of carcinoma. Lancet. 1902;159:1758–1761.
-
- Bernstein BE, Mikkelsen TS, Xie X, Kamal M, Huebert DJ, Cuff J, Fry B, Meissner A, Wernig M, Plath K, et al. A bivalent chromatin structure marks key developmental genes in embryonic stem cells. Cell. 2006;125:315–326. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P01 HL094374/HL/NHLBI NIH HHS/United States
- U01HL100405/HL/NHLBI NIH HHS/United States
- P01HL094374/HL/NHLBI NIH HHS/United States
- U01 HL100405/HL/NHLBI NIH HHS/United States
- U54HG004592/HG/NHGRI NIH HHS/United States
- U54HG007010/HG/NHGRI NIH HHS/United States
- P30DK056465/DK/NIDDK NIH HHS/United States
- P30 DK056465/DK/NIDDK NIH HHS/United States
- F30HL095343/HL/NHLBI NIH HHS/United States
- T32 GM007266/GM/NIGMS NIH HHS/United States
- F30 HL095343/HL/NHLBI NIH HHS/United States
- R01 HL084642/HL/NHLBI NIH HHS/United States
- F30 DK095678/DK/NIDDK NIH HHS/United States
- FDK095678A/PHS HHS/United States
- R01HL084642/HL/NHLBI NIH HHS/United States
- P01GM081719/GM/NIGMS NIH HHS/United States
- U01 ES017156/ES/NIEHS NIH HHS/United States
- P01 GM081619/GM/NIGMS NIH HHS/United States
- U54 HG007010/HG/NHGRI NIH HHS/United States
- U01ES017156/ES/NIEHS NIH HHS/United States
- U54 HG004592/HG/NHGRI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
