

Salmonella require the fatty acid regulator PPARδ for the establishment of a metabolic environment essential for long-term persistence
- PMID: 23954156
- PMCID: PMC3785333
- DOI: 10.1016/j.chom.2013.07.010
Salmonella require the fatty acid regulator PPARδ for the establishment of a metabolic environment essential for long-term persistence
Abstract
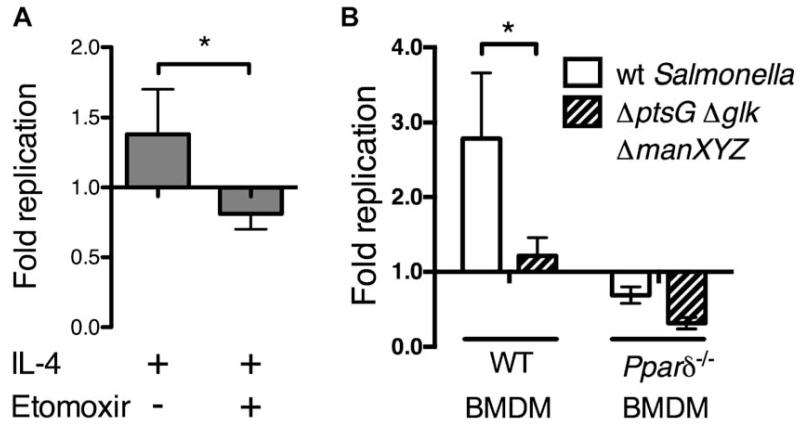
Host-adapted Salmonella strains are responsible for a number of disease manifestations in mammals, including an asymptomatic chronic infection in which bacteria survive within macrophages located in systemic sites. However, the host cell physiology and metabolic requirements supporting bacterial persistence are poorly understood. In a mouse model of long-term infection, we found that S. typhimurium preferentially associates with anti-inflammatory/M2 macrophages at later stages of infection. Further, PPARδ, a eukaryotic transcription factor involved in sustaining fatty acid metabolism, is upregulated in Salmonella-infected macrophages. PPARδ deficiency dramatically inhibits Salmonella replication, which is linked to the metabolic state of macrophages and the level of intracellular glucose available to bacteria. Pharmacological activation of PPARδ increases glucose availability and enhances bacterial replication in macrophages and mice, while Salmonella fail to persist in Pparδ null mice. These data suggest that M2 macrophages represent a unique niche for long-term intracellular bacterial survival and link the PPARδ-regulated metabolic state of the host cell to persistent bacterial infection.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures






Comment in
-
Bacterial persistence: finding the "sweet spot".Cell Host Microbe. 2013 Aug 14;14(2):119-20. doi: 10.1016/j.chom.2013.07.016. Cell Host Microbe. 2013. PMID: 23954150
References
Publication types
MeSH terms
Substances
Grants and funding
- DP1AR064158/AR/NIAMS NIH HHS/United States
- T32 AI007290/AI/NIAID NIH HHS/United States
- R01 AI089722/AI/NIAID NIH HHS/United States
- AI08972/AI/NIAID NIH HHS/United States
- HL076746/HL/NHLBI NIH HHS/United States
- R01 DK081405/DK/NIDDK NIH HHS/United States
- R01 HL076746/HL/NHLBI NIH HHS/United States
- T32 AI007328/AI/NIAID NIH HHS/United States
- DK081405/DK/NIDDK NIH HHS/United States
- T32 AI07290-28./AI/NIAID NIH HHS/United States
- DP1 AR064158/AR/NIAMS NIH HHS/United States
- T32 GM007276/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases