

Haemolysin coregulated protein is an exported receptor and chaperone of type VI secretion substrates
- PMID: 23954347
- PMCID: PMC3844553
- DOI: 10.1016/j.molcel.2013.07.025
Haemolysin coregulated protein is an exported receptor and chaperone of type VI secretion substrates
Abstract
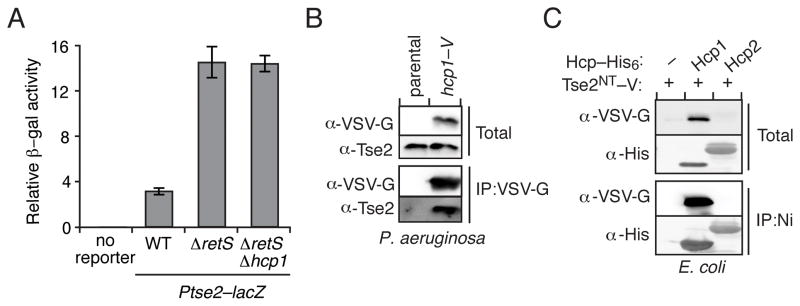
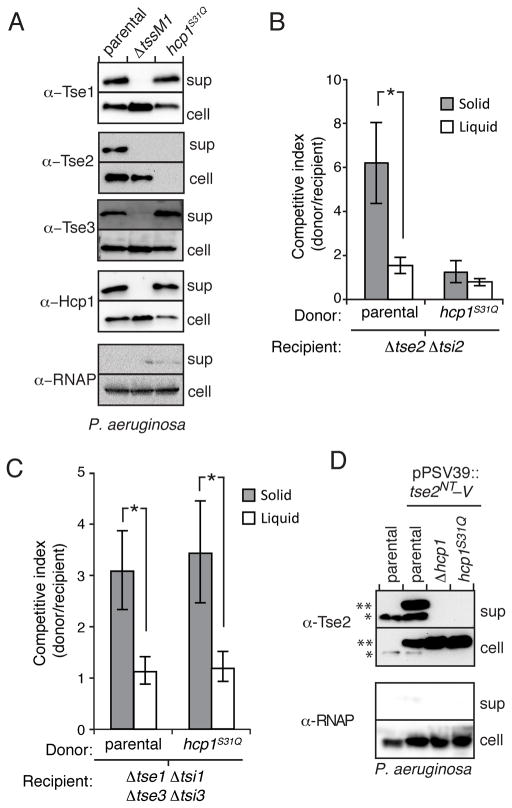
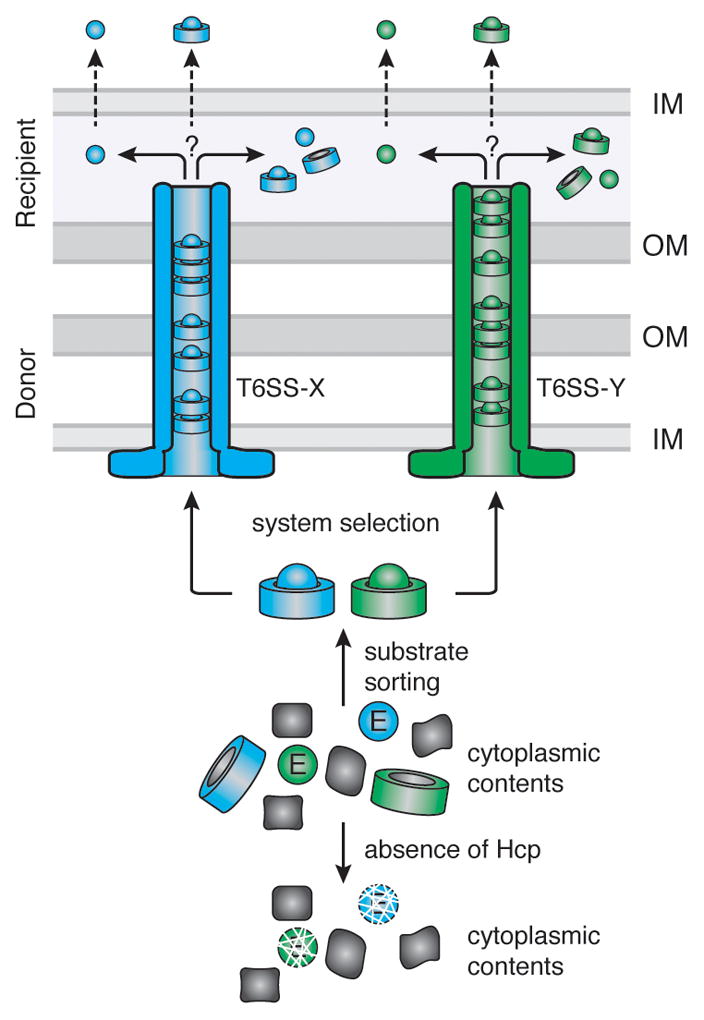
Secretion systems require high-fidelity mechanisms to discriminate substrates among the vast cytoplasmic pool of proteins. Factors mediating substrate recognition by the type VI secretion system (T6SS) of Gram-negative bacteria, a widespread pathway that translocates effector proteins into target bacterial cells, have not been defined. We report that haemolysin coregulated protein (Hcp), a ring-shaped hexamer secreted by all characterized T6SSs, binds specifically to cognate effector molecules. Electron microscopy analysis of an Hcp-effector complex from Pseudomonas aeruginosa revealed the effector bound to the inner surface of Hcp. Further studies demonstrated that interaction with the Hcp pore is a general requirement for secretion of diverse effectors encompassing several enzymatic classes. Though previous models depict Hcp as a static conduit, our data indicate it is a chaperone and receptor of substrates. These unique functions of a secreted protein highlight fundamental differences between the export mechanism of T6 and other characterized secretory pathways.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures




References
-
- Abdallah AM, Gey van Pittius NC, Champion PA, Cox J, Luirink J, Vandenbroucke-Grauls CM, Appelmelk BJ, Bitter W. Type VII secretion--mycobacteria show the way. Nat Rev Microbiol. 2007;5:883–891. - PubMed
-
- Akeda Y, Galan JE. Chaperone release and unfolding of substrates in type III secretion. Nature. 2005;437:911–915. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
