

Sphingolipids and lifespan regulation
- PMID: 23954556
- PMCID: PMC3925463
- DOI: 10.1016/j.bbalip.2013.08.006
Sphingolipids and lifespan regulation
Abstract
Diseases including cancer, type 2 diabetes, cardiovascular and immune dysfunction and neurodegeneration become more prevalent as we age, and combined with the increase in average human lifespan, place an ever increasing burden on the health care system. In this chapter we focus on finding ways of modulating sphingolipids to prevent the development of age-associated diseases or delay their onset, both of which could improve health in elderly, fragile people. Reducing the incidence of or delaying the onset of diseases of aging has blossomed in the past decade because of advances in understanding signal transduction pathways and cellular processes, especially in model organisms, that are largely conserved in most eukaryotes and that can be modulated to reduce signs of aging and increase health span. In model organisms such interventions must also increase lifespan to be considered significant, but this is not a requirement for use in humans. The most encouraging interventions in model organisms involve lowering the concentration of one or more sphingolipids so as to reduce the activity of key signaling pathways, one of the most promising being the Target of Rapamycin Complex 1 (TORC1) protein kinase pathway. Other potential ways in which modulating sphingolipids may contribute to improving the health profile of the elderly is by reducing oxidative stresses, inflammatory responses and growth factor signaling. Lastly, perhaps the most interesting way to modulate sphingolipids and promote longevity is by lowering the activity of serine palmitoyltransferase, the first enzyme in the de novo sphingolipid biosynthesis pathway. Available data in yeasts and rodents are encouraging and as we gain insights into molecular mechanisms the strategies for improving human health by modulating sphingolipids will become more apparent. This article is part of a Special Issue entitled New Frontiers in Sphingolipid Biology.
Keywords: Aging; Autophagy; Ceramide; Myriocin; Sphingosing-1-phosphate.
© 2013.
Figures
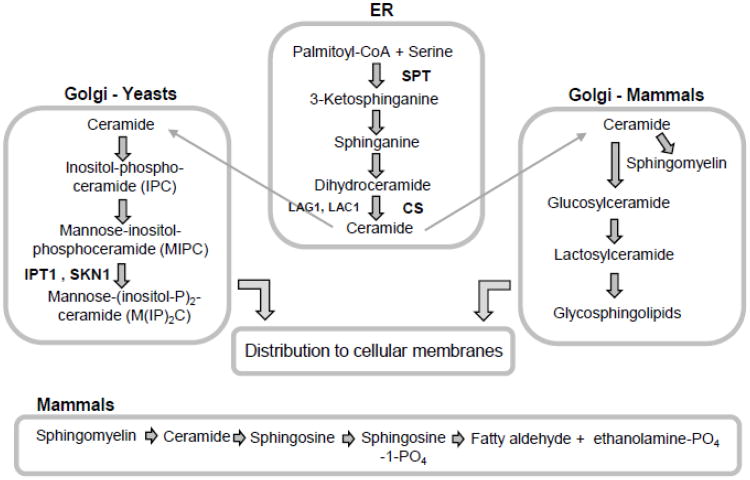
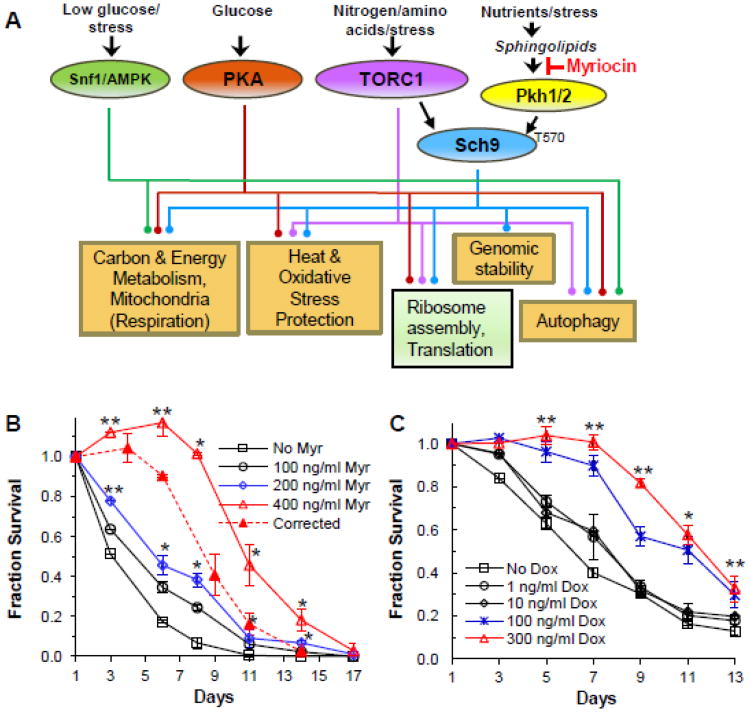
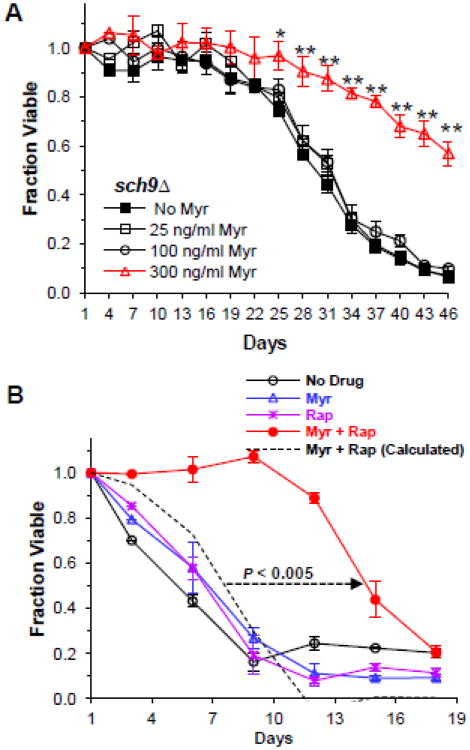
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
