

Impact of environmental conditions on the form and function of Candida albicans biofilms
- PMID: 23954841
- PMCID: PMC3811334
- DOI: 10.1128/EC.00127-13
Impact of environmental conditions on the form and function of Candida albicans biofilms
Abstract
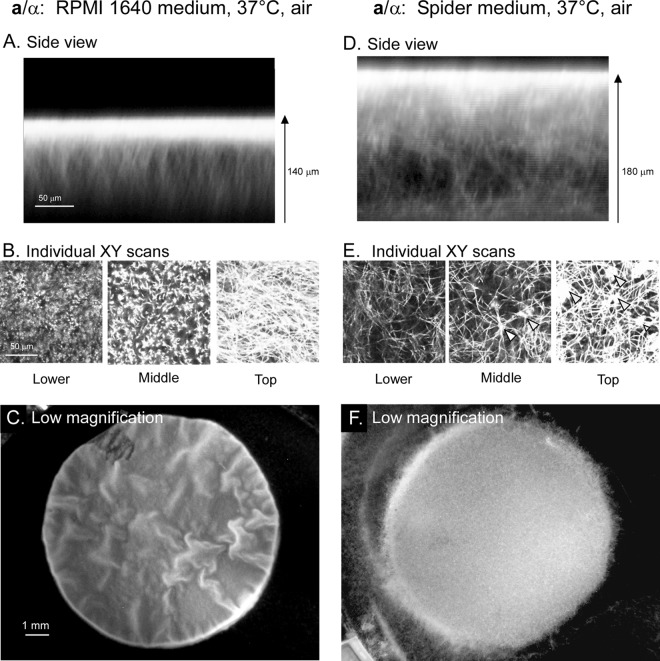
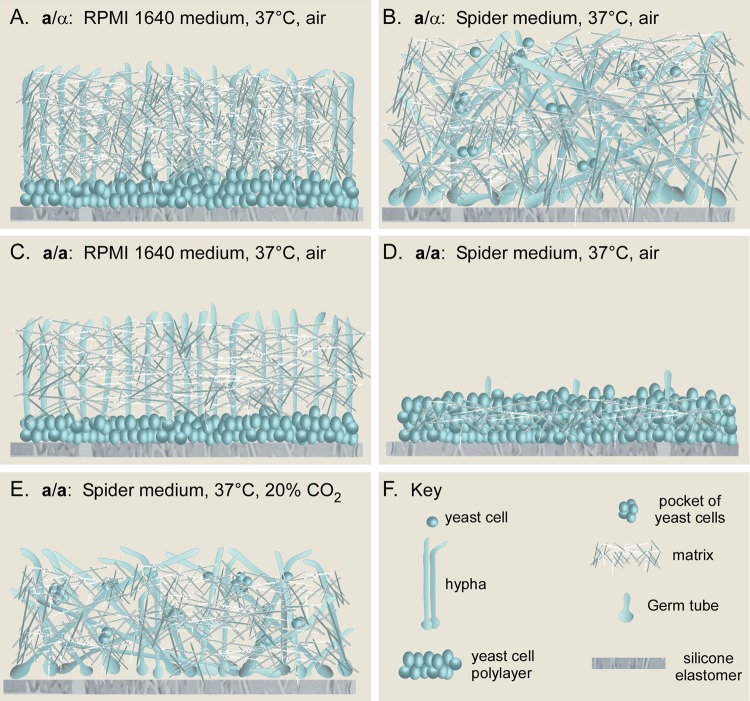
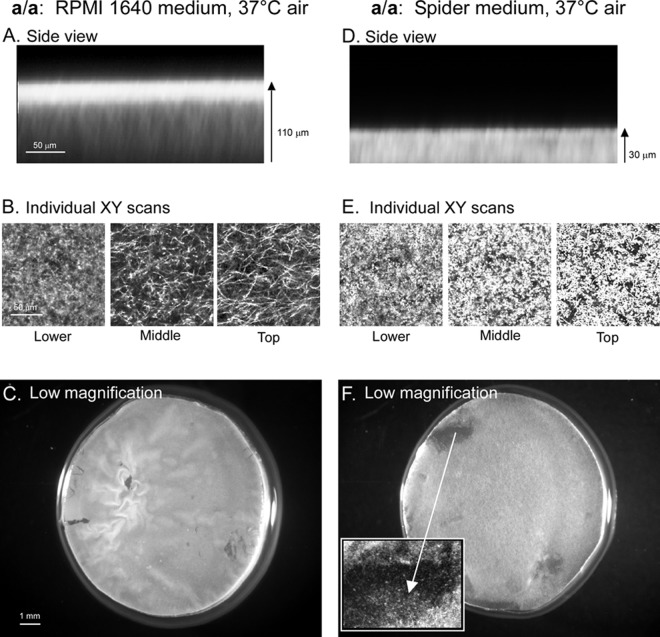
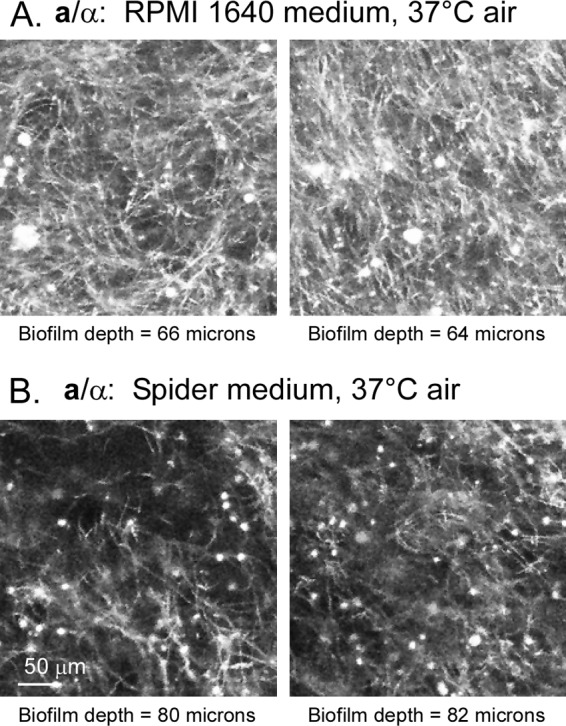
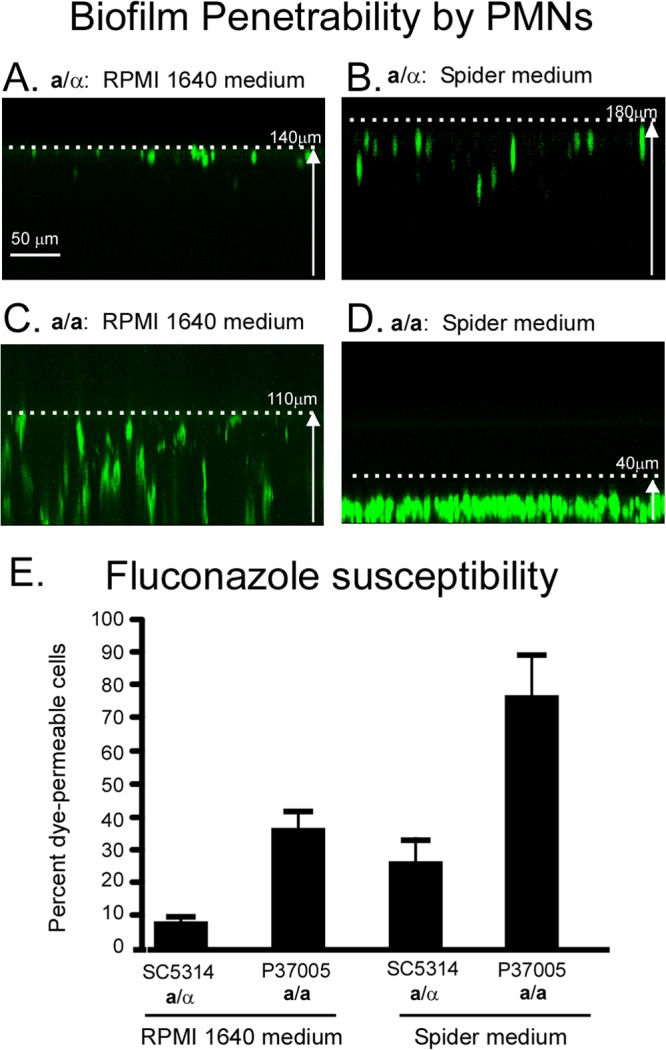
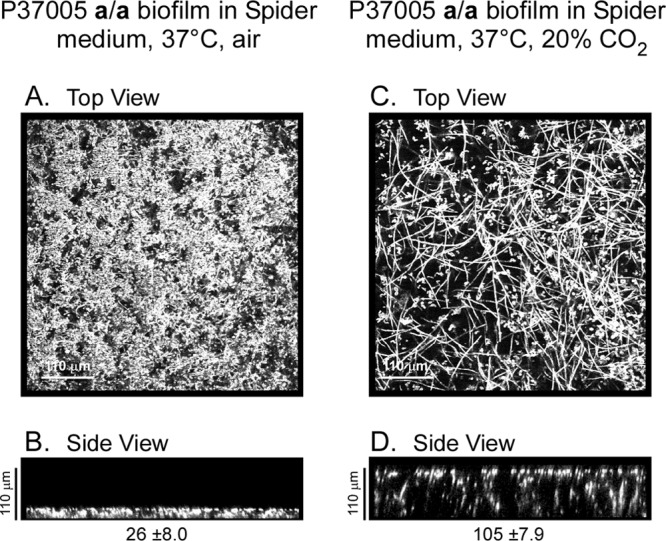
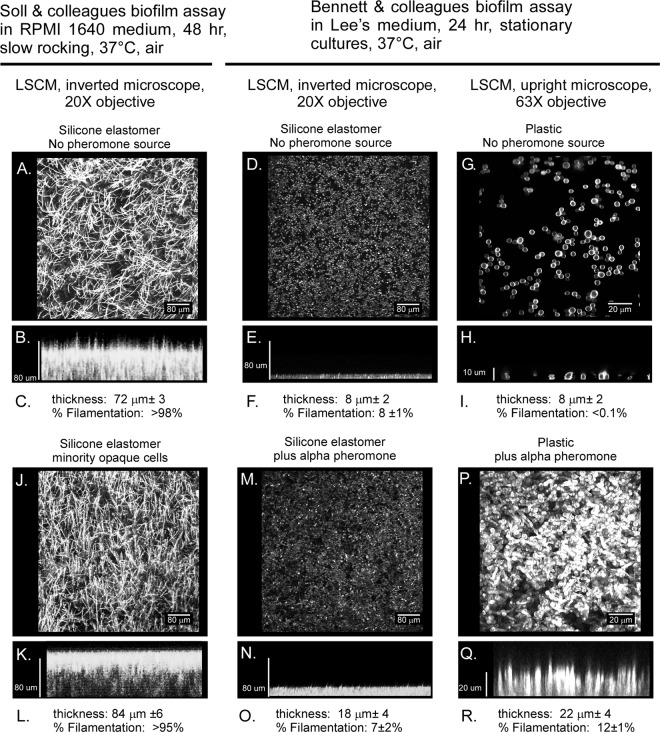
Candida albicans, like other pathogens, can form complex biofilms on a variety of substrates. However, as the number of studies of gene regulation, architecture, and pathogenic traits of C. albicans biofilms has increased, so have differences in results. This suggests that depending upon the conditions employed, biofilms may vary widely, thus hampering attempts at a uniform description. Gene expression studies suggest that this may be the case. To explore this hypothesis further, we compared the architectures and traits of biofilms formed in RPMI 1640 and Spider media at 37°C in air. Biofilms formed by a/α cells in the two media differed to various degrees in cellular architecture, matrix deposition, penetrability by leukocytes, fluconazole susceptibility, and the facilitation of mating. Similar comparisons of a/a cells in the two media, however, were made difficult given that in air, although a/a cells form traditional biofilms in RPMI medium, they form polylayers composed primarily of yeast cells in Spider medium. These polylayers lack an upper hyphal/matrix region, are readily penetrated by leukocytes, are highly fluconazole susceptible, and do not facilitate mating. If, however, air is replaced with 20% CO2, a/a cells make a biofilm in Spider medium similar architecturally to that of a/α cells, which facilitates mating. A second, more cursory comparison is made between the disparate cellular architectures of a/a biofilms formed in air in RPMI and Lee's media. The results demonstrate that C. albicans forms very different types of biofilms depending upon the composition of the medium, level of CO2 in the atmosphere, and configuration of the MTL locus.
Figures
References
-
- Costerton JW, Stewart PS, Greenberg EP. 1999. Bacterial biofilms: a common cause of persistent infections. Science 284:1318–1322 - PubMed
-
- Hall-Stoodley L, Stoodley P. 2005. Biofilm formation and dispersal and the transmission of human pathogens. Trends Microbiol. 13:7–10 - PubMed
-
- Douglas LJ. 2003. Candida biofilms and their role in infection. Trends Microbiol. 11:30–36 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
