

Ovariectomy expands murine short-term hemopoietic stem cell function through T cell expressed CD40L and Wnt10B
- PMID: 23954891
- PMCID: PMC3790505
- DOI: 10.1182/blood-2013-03-487801
Ovariectomy expands murine short-term hemopoietic stem cell function through T cell expressed CD40L and Wnt10B
Abstract
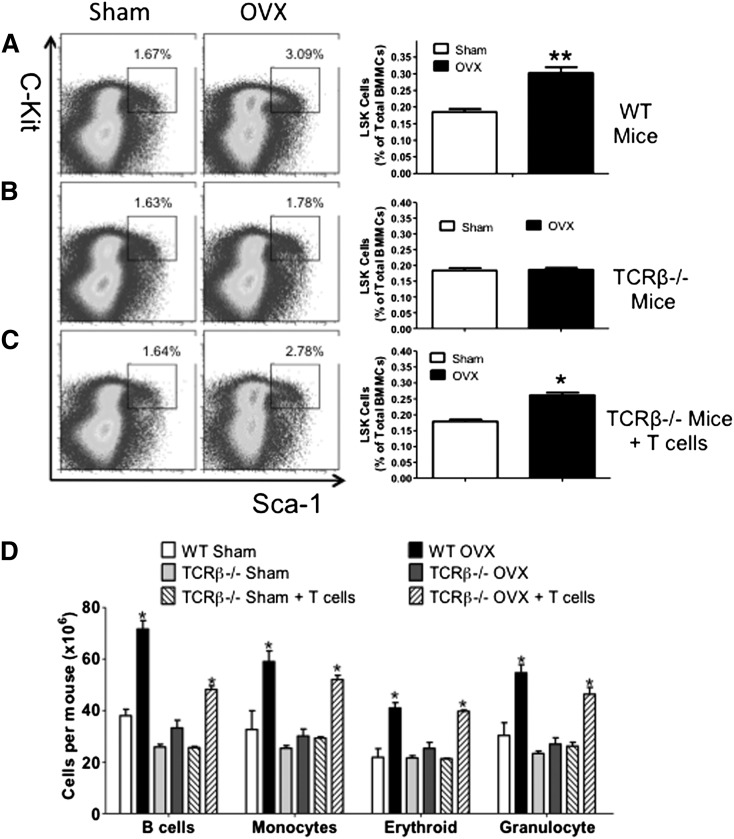
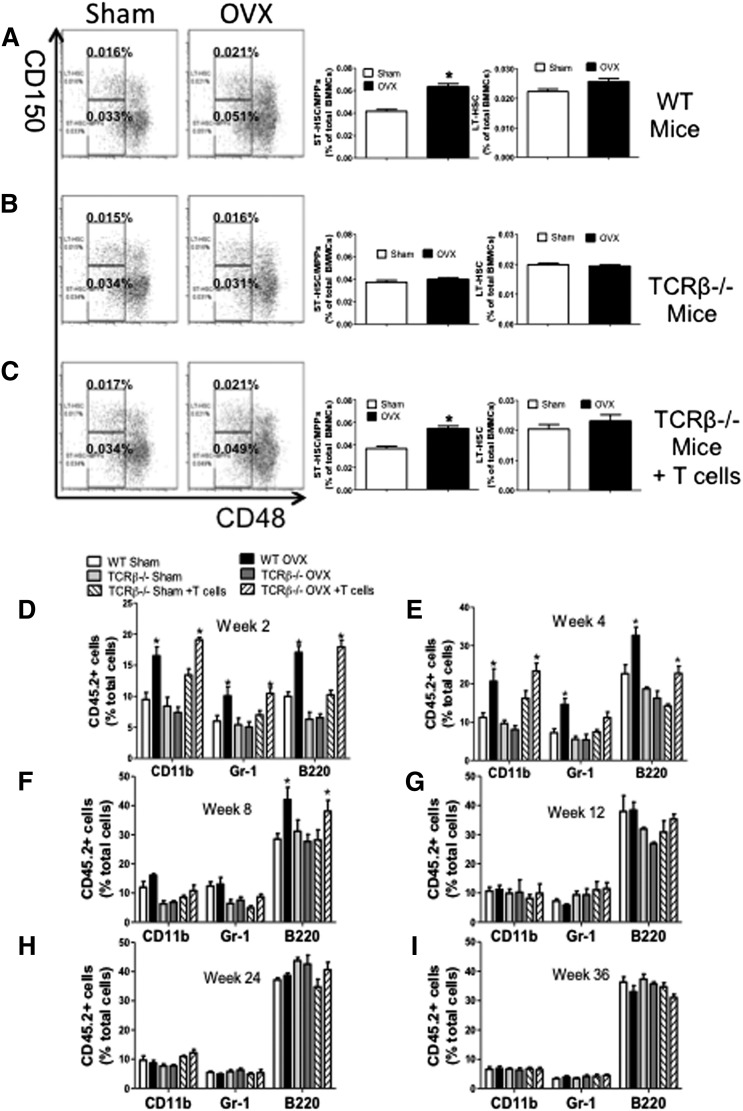
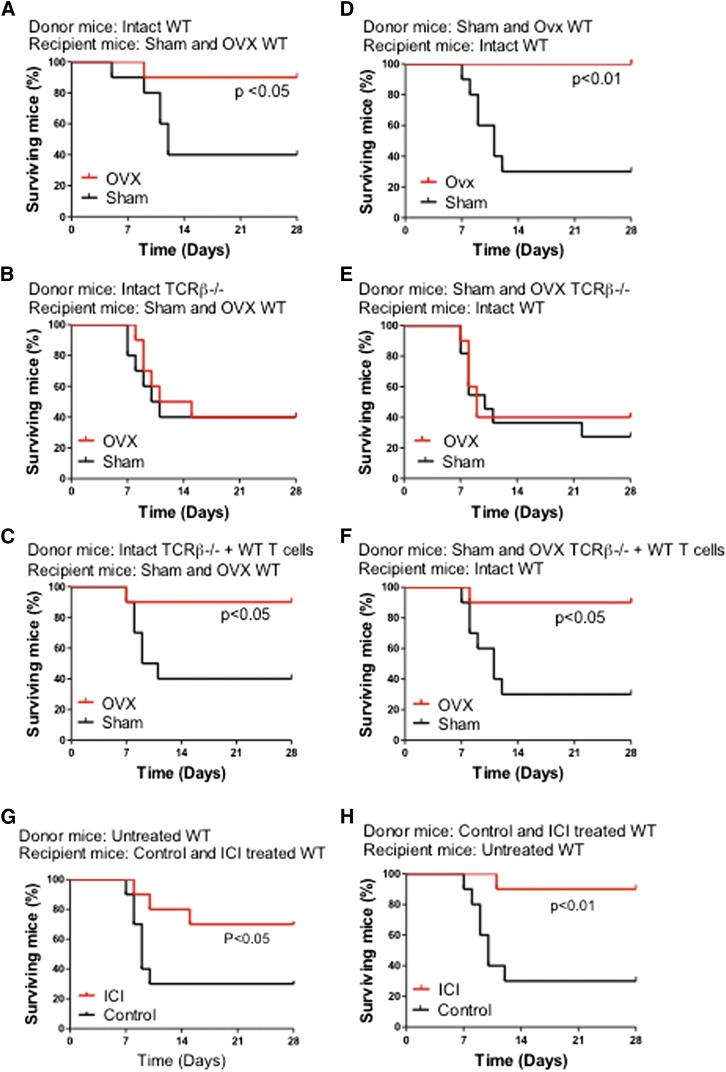
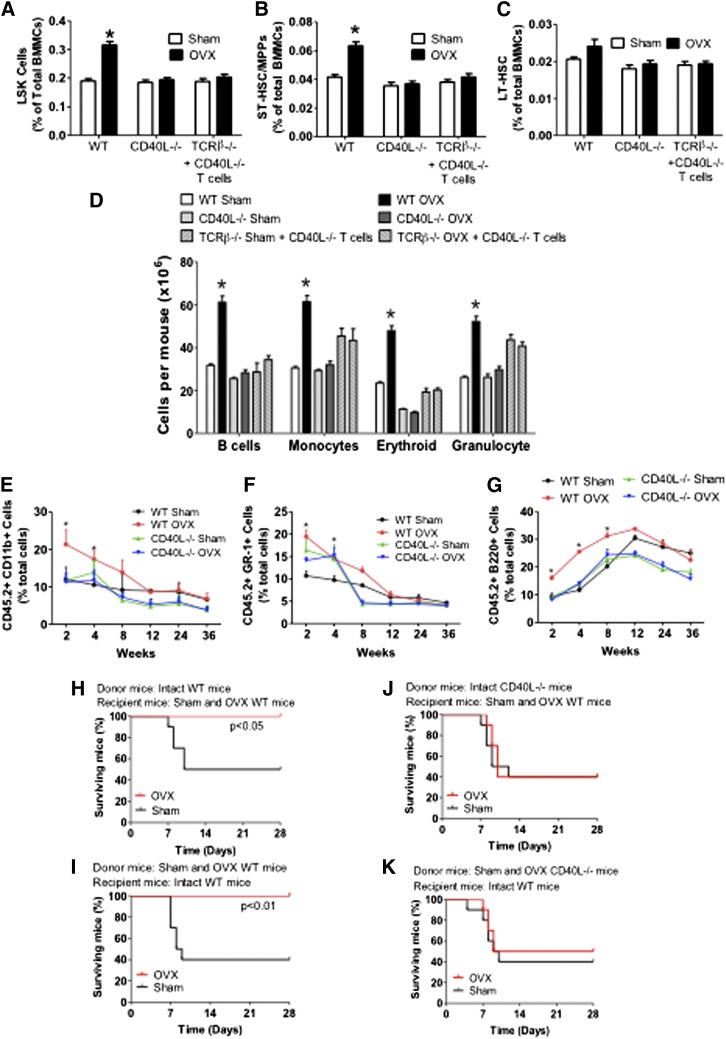
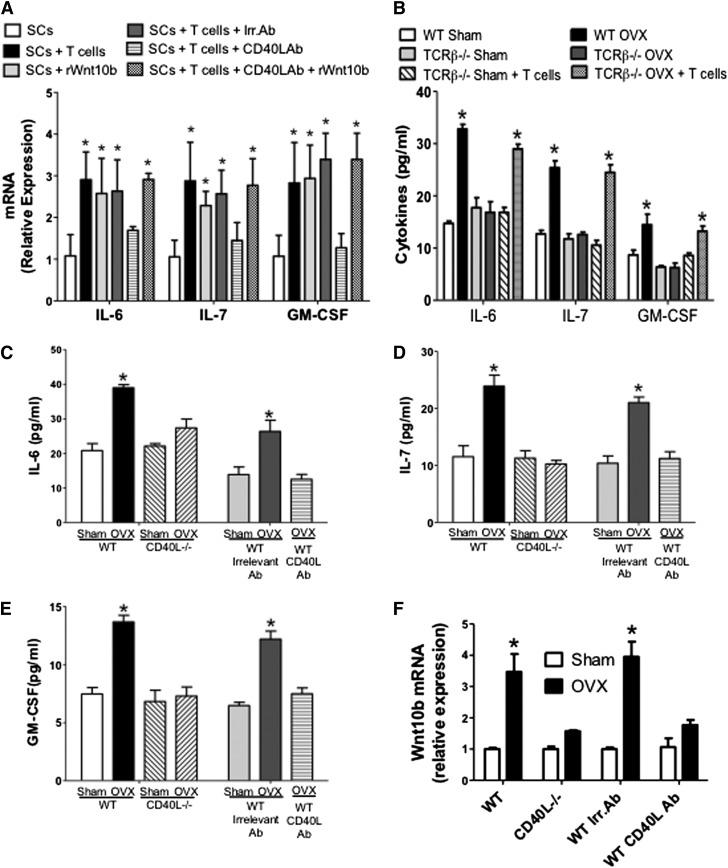
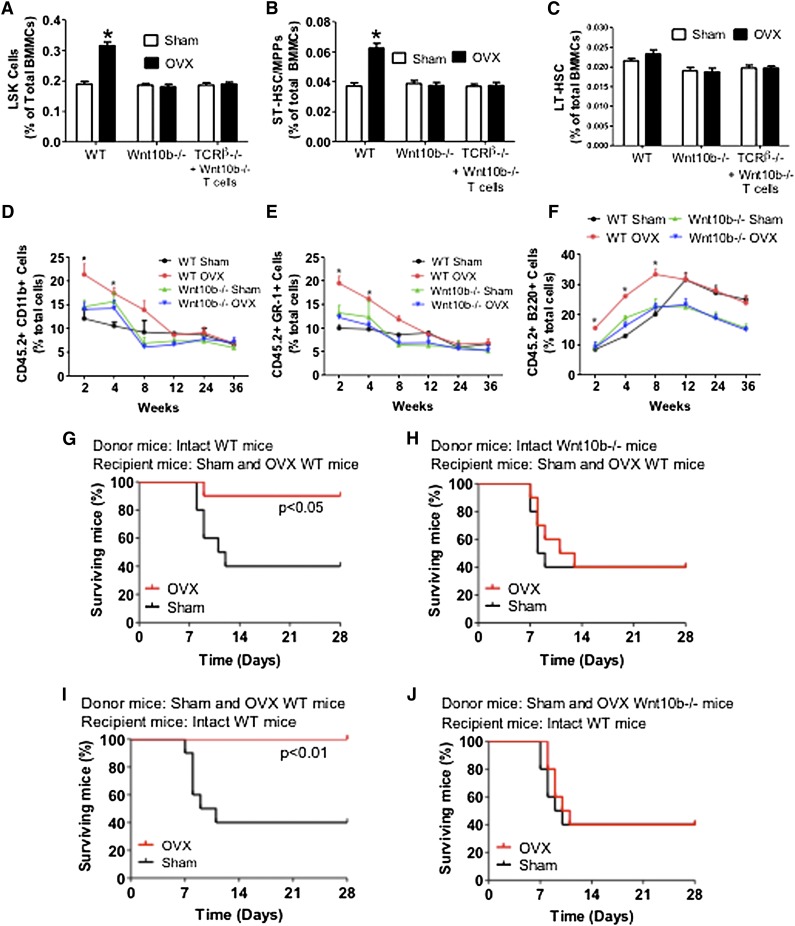
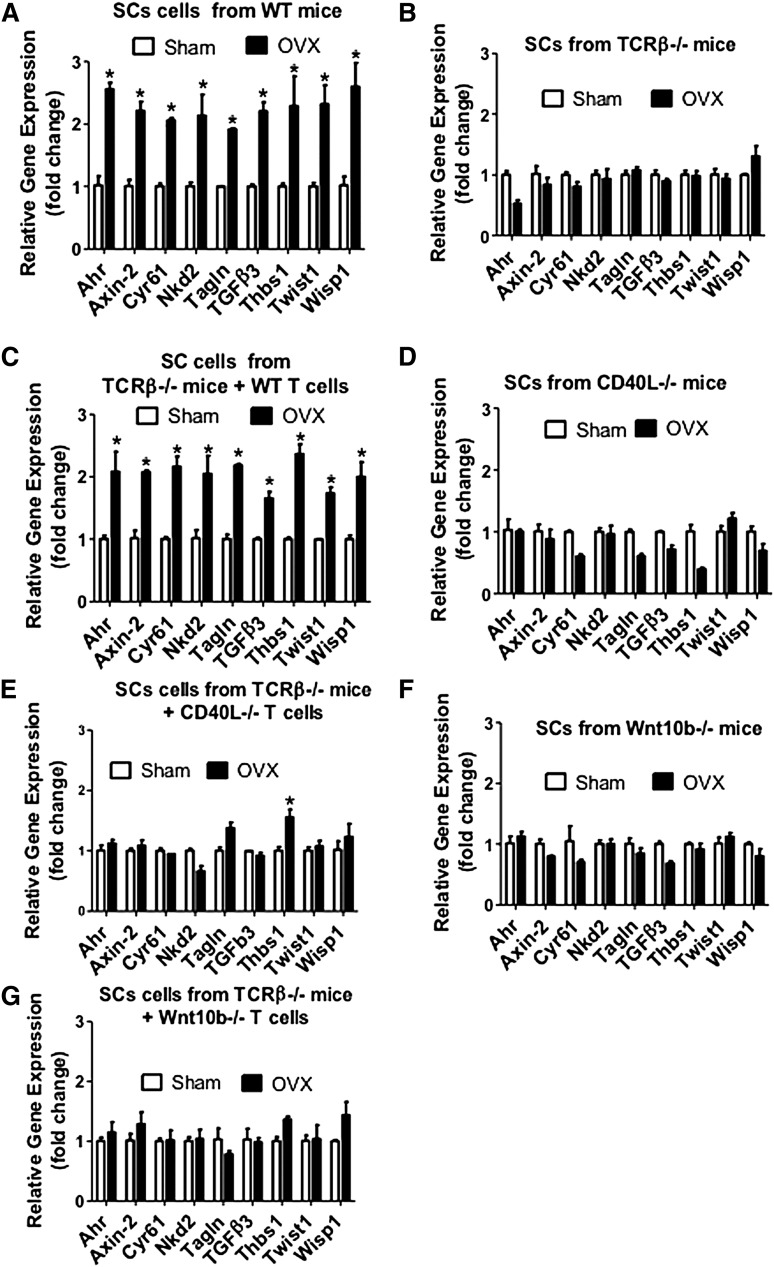
Estrogen deficiency expands hemopoietic stem and progenitor cells (HSPCs) and mature blood lineages, but the involved mechanism and the affected HSPC populations are mostly unknown. Here we show that ovariectomy (ovx) expands short-term HSPCs (ST-HSPCs) and improves blood cell engraftment and host survival after bone marrow (BM) transplantation through a dual role of the T-cell costimulatory molecule CD40 ligand (CD40L). This surface receptor is required for ovx to stimulate T-cell production of Wnt10b, a Wnt ligand that activates Wnt signaling in HSPCs and stromal cells (SCs). Moreover, CD40L is required for ovx to increase SC production of the hemopoietic cytokines interleukin (IL)-6, IL-7, and granulocyte macrophage-colony-stimulating factor. Attesting to the relevance of CD40L and Wnt10b, ovx fails to expand ST-HSPCs in CD40L-null mice and in animals lacking global or T-cell expression of Wnt10b. In summary, T cells expressed CD40L, and the resulting increased production of Wnt10b and hemopoietic cytokines by T cells and SCs, respectively, plays a pivotal role in the mechanism by which ovx regulates hemopoiesis. The data suggest that antiestrogens may represent pharmacological targets to improve ST-HSPC function through activation of the microenvironment.
Figures
Similar articles
-
PTH expands short-term murine hemopoietic stem cells through T cells.Blood. 2012 Nov 22;120(22):4352-62. doi: 10.1182/blood-2012-06-438531. Epub 2012 Sep 5. Blood. 2012. PMID: 22955916 Free PMC article.
-
T cell-expressed CD40L potentiates the bone anabolic activity of intermittent PTH treatment.J Bone Miner Res. 2015 Apr;30(4):695-705. doi: 10.1002/jbmr.2394. J Bone Miner Res. 2015. PMID: 25359628 Free PMC article.
-
Ovariectomy disregulates osteoblast and osteoclast formation through the T-cell receptor CD40 ligand.Proc Natl Acad Sci U S A. 2011 Jan 11;108(2):768-73. doi: 10.1073/pnas.1013492108. Epub 2010 Dec 27. Proc Natl Acad Sci U S A. 2011. PMID: 21187391 Free PMC article.
-
Osteoimmunology and its implications for transplantation.Am J Transplant. 2013 Sep;13(9):2245-54. doi: 10.1111/ajt.12380. Epub 2013 Aug 5. Am J Transplant. 2013. PMID: 23915249 Review.
-
Parallels between immune driven-hematopoiesis and T cell activation: 3 signals that relay inflammatory stress to the bone marrow.Exp Cell Res. 2014 Dec 10;329(2):239-47. doi: 10.1016/j.yexcr.2014.09.016. Epub 2014 Sep 22. Exp Cell Res. 2014. PMID: 25246130 Review.
Cited by
-
Hydrogen Sulfide Is a Novel Regulator of Bone Formation Implicated in the Bone Loss Induced by Estrogen Deficiency.J Bone Miner Res. 2016 May;31(5):949-63. doi: 10.1002/jbmr.2757. Epub 2015 Dec 23. J Bone Miner Res. 2016. PMID: 26614970 Free PMC article.
-
Ovariectomy induces bone loss via microbial-dependent trafficking of intestinal TNF+ T cells and Th17 cells.J Clin Invest. 2021 Feb 15;131(4):e143137. doi: 10.1172/JCI143137. J Clin Invest. 2021. PMID: 33586672 Free PMC article.
-
Biology of BM failure syndromes: role of microenvironment and niches.Hematology Am Soc Hematol Educ Program. 2014 Dec 5;2014(1):71-6. doi: 10.1182/asheducation-2014.1.71. Epub 2014 Nov 18. Hematology Am Soc Hematol Educ Program. 2014. PMID: 25696837 Free PMC article. Review.
-
Luteinizing hormone signaling restricts hematopoietic stem cell expansion during puberty.EMBO J. 2018 Sep 3;37(17):e98984. doi: 10.15252/embj.201898984. Epub 2018 Jul 23. EMBO J. 2018. PMID: 30037826 Free PMC article.
-
Cyclic Adenosine Monophosphate (cAMP)-Dependent Phosphodiesterase Inhibition Promotes Bone Anabolism Through CD8+ T Cell Wnt-10b Production in Mice.JBMR Plus. 2022 May 31;6(7):e10636. doi: 10.1002/jbm4.10636. eCollection 2022 Jul. JBMR Plus. 2022. PMID: 35866149 Free PMC article.
References
-
- Thurmond TS, Murante FG, Staples JE, Silverstone AE, Korach KS, Gasiewicz TA. Role of estrogen receptor alpha in hematopoietic stem cell development and B lymphocyte maturation in the male mouse. Endocrinology. 2000;141(7):2309–2318. - PubMed
-
- Zoller AL, Kersh GJ. Estrogen induces thymic atrophy by eliminating early thymic progenitors and inhibiting proliferation of beta-selected thymocytes. J Immunol. 2006;176(12):7371–7378. - PubMed
-
- Jilka RL, Passeri G, Girasole G, et al. Estrogen loss upregulates hematopoiesis in the mouse: a mediating role of IL-6. Exp Hematol. 1995;23(6):500–506. - PubMed
-
- Erben RG, Raith S, Eberle J, Stangassinger M. Ovariectomy augments B lymphopoiesis and generation of monocyte-macrophage precursors in rat bone marrow. Am J Physiol. 1998;274(3 Pt 1):E476–E483. - PubMed
-
- Riggs BL, Khosla S, Melton LJ., III Sex steroids and the construction and conservation of the adult skeleton. Endocr Rev. 2002;23(3):279–302. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- AR053607/AR/NIAMS NIH HHS/United States
- R01 DK091780/DK/NIDDK NIH HHS/United States
- K01 AR061453/AR/NIAMS NIH HHS/United States
- AR056090/AR/NIAMS NIH HHS/United States
- R01 AR056090/AR/NIAMS NIH HHS/United States
- AR49659/AR/NIAMS NIH HHS/United States
- AR059364/AR/NIAMS NIH HHS/United States
- AG040013/AG/NIA NIH HHS/United States
- I01 BX000105/BX/BLRD VA/United States
- AR061453/AR/NIAMS NIH HHS/United States
- R01 AG040013/AG/NIA NIH HHS/United States
- R01 AR049659/AR/NIAMS NIH HHS/United States
- R21 AR053607/AR/NIAMS NIH HHS/United States
- R01 AR059364/AR/NIAMS NIH HHS/United States
- DK091780/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
