

Improving the lethal effect of cpl-7, a pneumococcal phage lysozyme with broad bactericidal activity, by inverting the net charge of its cell wall-binding module
- PMID: 23959317
- PMCID: PMC3811316
- DOI: 10.1128/AAC.01372-13
Improving the lethal effect of cpl-7, a pneumococcal phage lysozyme with broad bactericidal activity, by inverting the net charge of its cell wall-binding module
Erratum in
- Antimicrob Agents Chemother. 2014 May;58(5):2996. de Paz, Héctor [corrected to de Paz, Héctor D]
Abstract
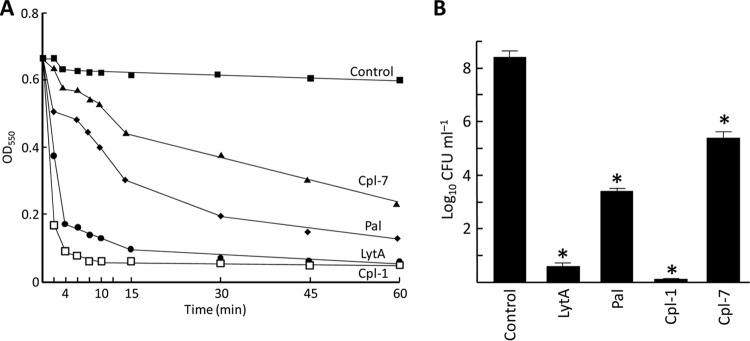
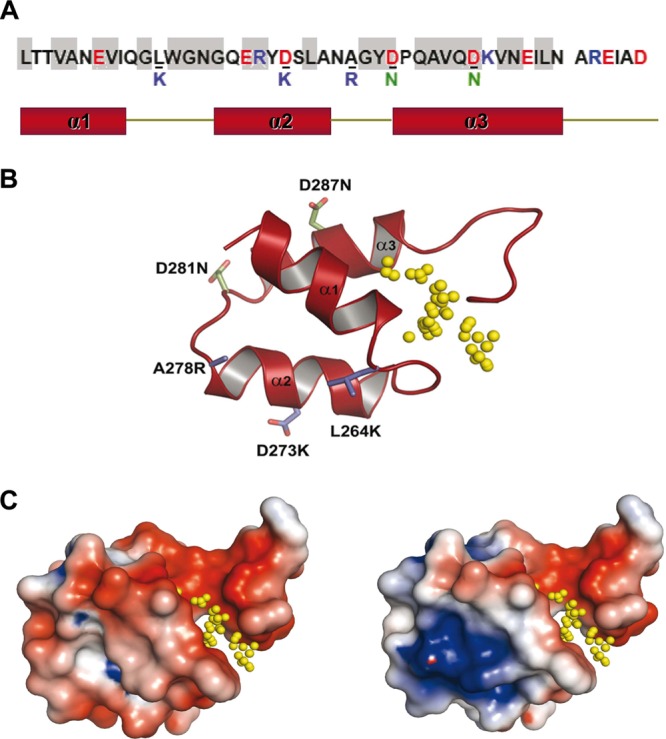
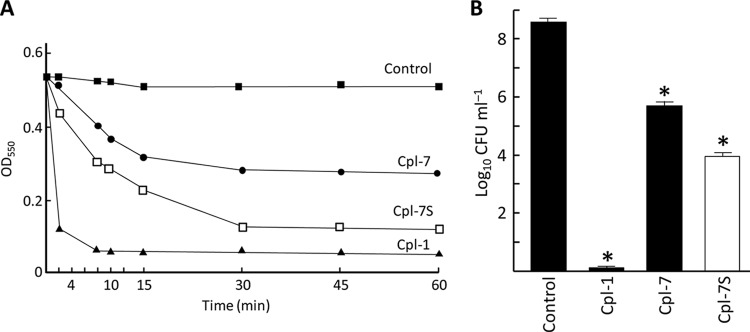
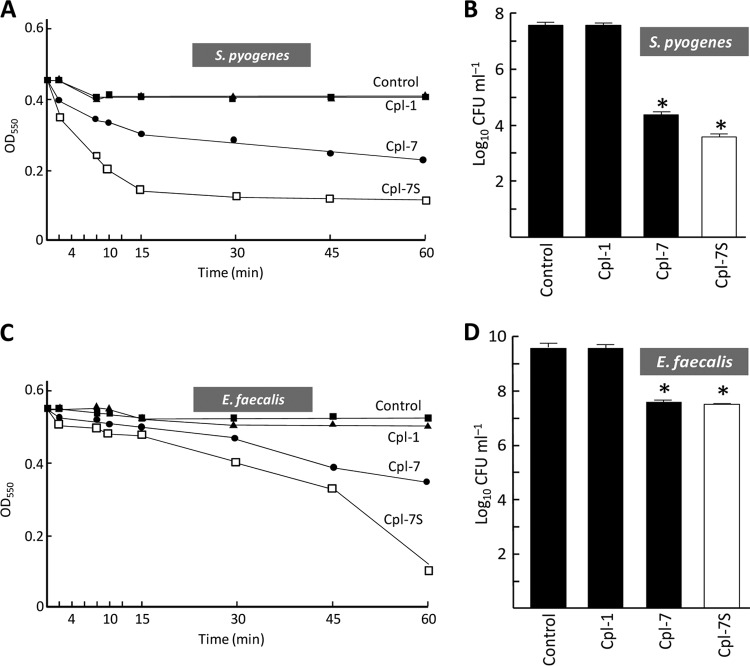
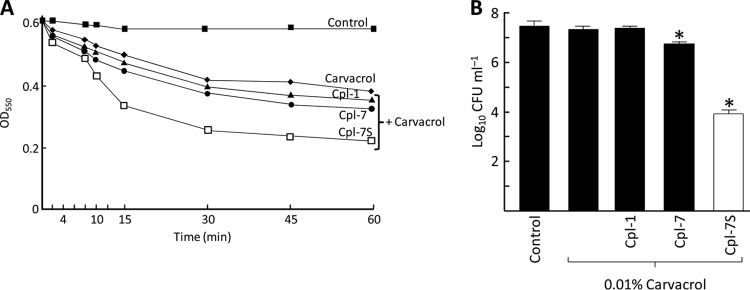
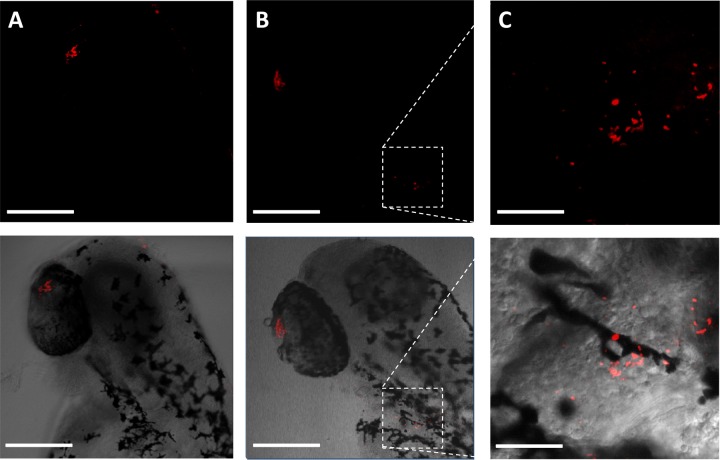
Phage endolysins are murein hydrolases that break the bacterial cell wall to provoke lysis and release of phage progeny. Recently, these enzymes have also been recognized as powerful and specific antibacterial agents when added exogenously. In the pneumococcal system, most cell wall associated murein hydrolases reported so far depend on choline for activity, and Cpl-7 lysozyme constitutes a remarkable exception. Here, we report the improvement of the killing activity of the Cpl-7 endolysin by inversion of the sign of the charge of the cell wall-binding module (from -14.93 to +3.0 at neutral pH). The engineered variant, Cpl-7S, has 15 amino acid substitutions and an improved lytic activity against Streptococcus pneumoniae (including multiresistant strains), Streptococcus pyogenes, and other pathogens. Moreover, we have demonstrated that a single 25-μg dose of Cpl-7S significantly increased the survival rate of zebrafish embryos infected with S. pneumoniae or S. pyogenes, confirming the killing effect of Cpl-7S in vivo. Interestingly, Cpl-7S, in combination with 0.01% carvacrol (an essential oil), was also found to efficiently kill Gram-negative bacteria such as Escherichia coli and Pseudomonas putida, an effect not described previously. Our findings provide a strategy to improve the lytic activity of phage endolysins based on facilitating their pass through the negatively charged bacterial envelope, and thereby their interaction with the cell wall target, by modulating the net charge of the cell wall-binding modules.
Figures

References
-
- Maestro B, Sanz JM. 2007. Novel approaches to fight Streptococcus pneumoniae. Recent Pat. Antiinfect. Drug Discov. 2:188–196 - PubMed
-
- WHO 2007. Pneumococcal conjugate vaccine for childhood immunization—WHO position paper. Wkly. Epidemiol. Rec. 82:93–104 - PubMed
-
- Pittet LF, Posfay-Barbe KM. 2012. Pneumococcal vaccines for children: a global public health priority. Clin. Microbiol. Infect. 18(Suppl 5):25–36 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
