

Metal switch-controlled myosin II from Dictyostelium discoideum supports closure of nucleotide pocket during ATP binding coupled to detachment from actin filaments
- PMID: 23960071
- PMCID: PMC3784747
- DOI: 10.1074/jbc.M113.466045
Metal switch-controlled myosin II from Dictyostelium discoideum supports closure of nucleotide pocket during ATP binding coupled to detachment from actin filaments
Abstract
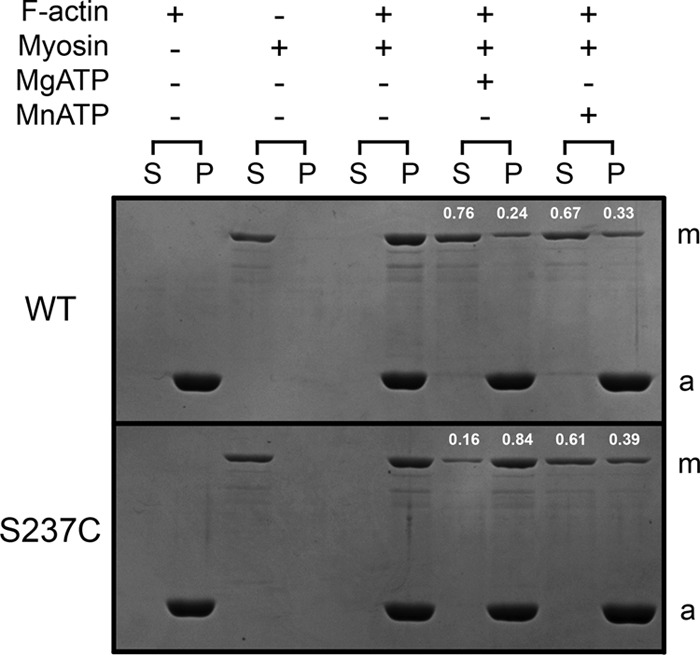
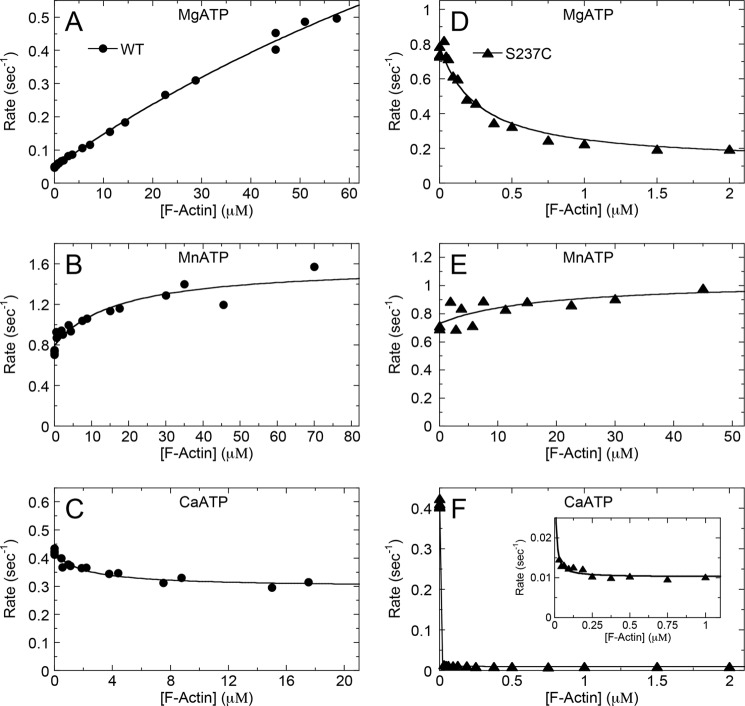
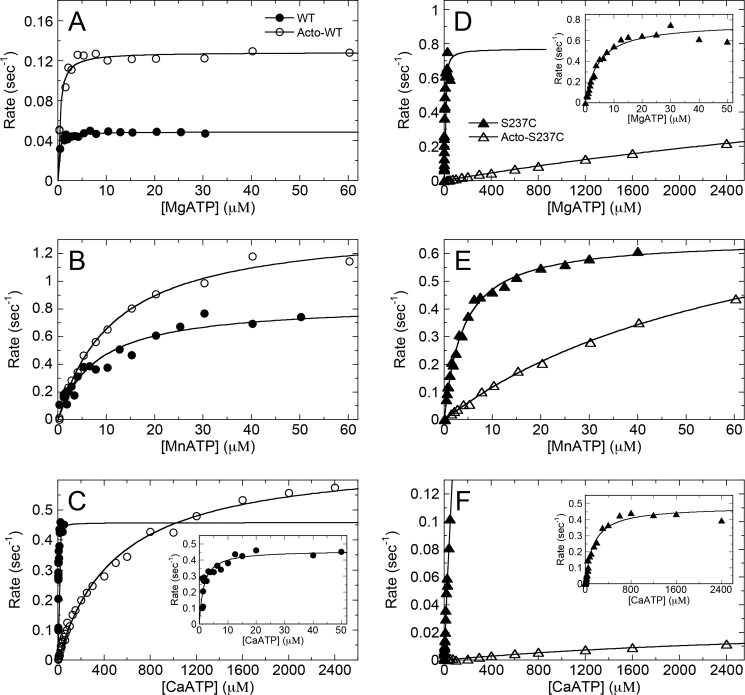
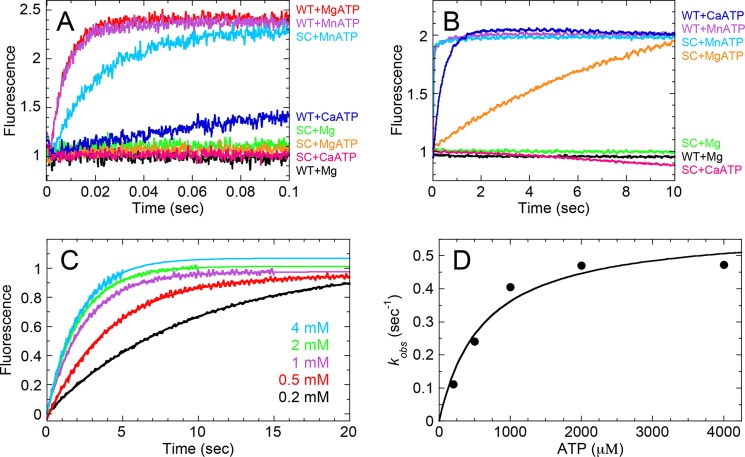
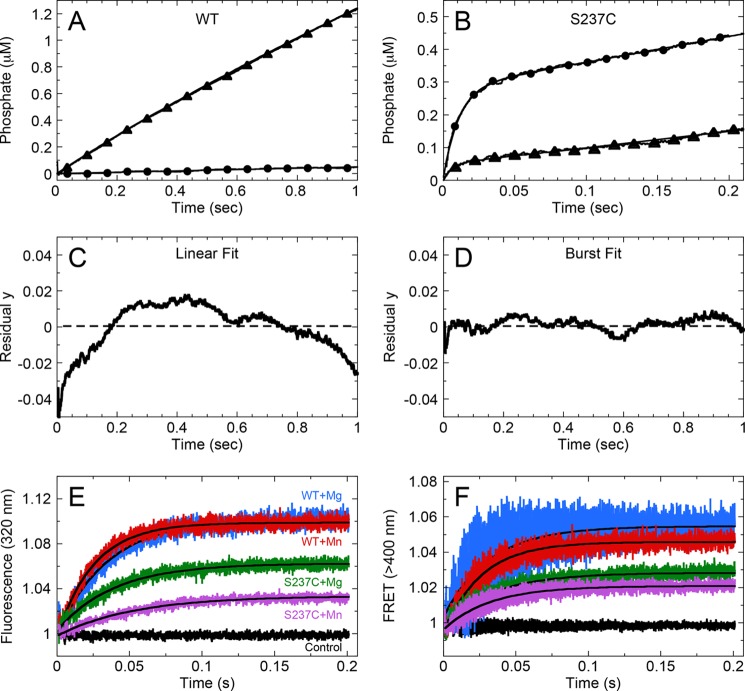
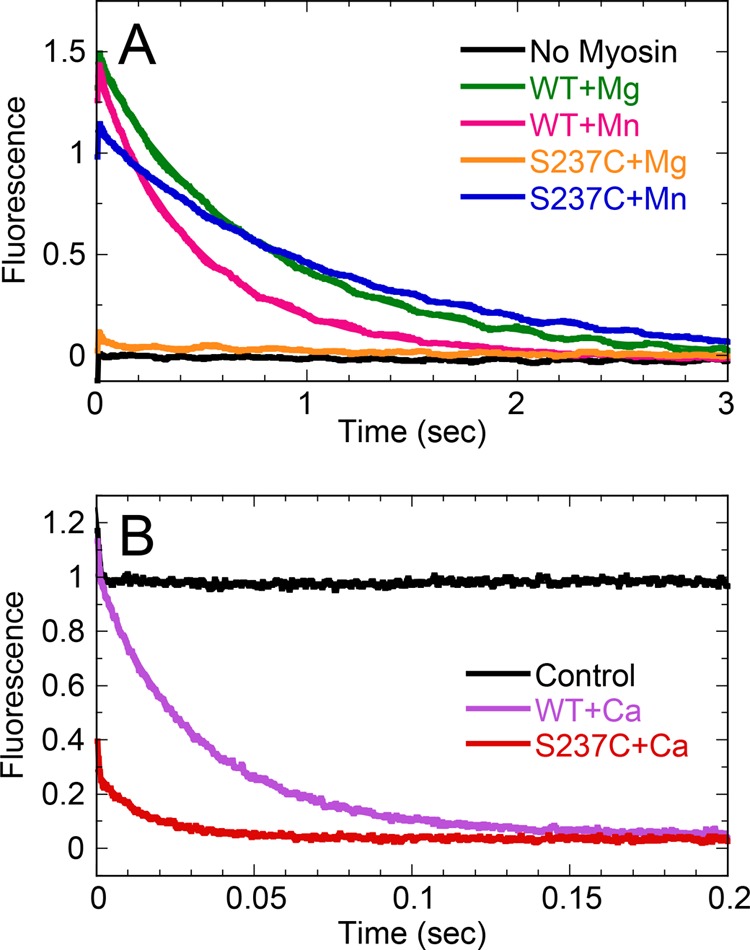
G-proteins, kinesins, and myosins are hydrolases that utilize a common protein fold and divalent metal cofactor (typically Mg(2+)) to coordinate purine nucleotide hydrolysis. The nucleoside triphosphorylase activities of these enzymes are activated through allosteric communication between the nucleotide-binding site and the activator/effector/polymer interface to convert the free energy of nucleotide hydrolysis into molecular switching (G-proteins) or force generation (kinesins and myosin). We have investigated the ATPase mechanisms of wild-type and the S237C mutant of non-muscle myosin II motor from Dictyostelium discoideum. The S237C substitution occurs in the conserved metal-interacting switch-1, and we show that this substitution modulates the actomyosin interaction based on the divalent metal present in solution. Surprisingly, S237C shows rapid basal steady-state Mg(2+)- or Mn(2+)-ATPase kinetics, but upon binding actin, its MgATPase is inhibited. This actin inhibition is relieved by Mn(2+), providing a direct and experimentally reversible linkage of switch-1 and the actin-binding cleft through the swapping of divalent metals in the reaction. Using pyrenyl-labeled F-actin, we demonstrate that acto·S237C undergoes slow and weak MgATP binding, which limits the rate of steady-state catalysis. Mn(2+) rescues this effect to near wild-type activity. 2'(3')-O-(N-Methylanthraniloyl)-ADP release experiments show the need for switch-1 interaction with the metal cofactor for tight ADP binding. Our results are consistent with strong reciprocal coupling of nucleoside triphosphate and F-actin binding and provide additional evidence for the allosteric communication pathway between the nucleotide-binding site and the filament-binding region.
Keywords: ATPases; Actin; Manganese; Molecular Motors; Myosin.
Figures

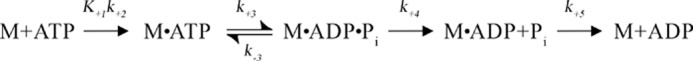
Similar articles
-
The Conserved Lysine-265 Allosterically Modulates Nucleotide- and Actin-binding Site Coupling in Myosin-2.Sci Rep. 2017 Aug 9;7(1):7650. doi: 10.1038/s41598-017-07933-y. Sci Rep. 2017. PMID: 28794442 Free PMC article.
-
The effect of F-actin on the relay helix position of myosin II, as revealed by tryptophan fluorescence, and its implications for mechanochemical coupling.Biochemistry. 2004 Dec 14;43(49):15404-17. doi: 10.1021/bi048338j. Biochemistry. 2004. PMID: 15581352
-
Allosteric communication in Dictyostelium myosin II.J Muscle Res Cell Motil. 2012 Oct;33(5):305-12. doi: 10.1007/s10974-012-9304-y. Epub 2012 Jul 1. J Muscle Res Cell Motil. 2012. PMID: 22752265 Free PMC article.
-
Actin polymerization: regulation by divalent metal ion and nucleotide binding, ATP hydrolysis and binding of myosin.Adv Exp Med Biol. 1994;358:71-81. doi: 10.1007/978-1-4615-2578-3_7. Adv Exp Med Biol. 1994. PMID: 7801813 Review.
-
Dictyostelium myosin II as a model to study the actin-myosin interactions during force generation.J Muscle Res Cell Motil. 2002;23(7-8):697-702. doi: 10.1023/a:1024415409406. J Muscle Res Cell Motil. 2002. PMID: 12952068 Review.
Cited by
-
Direct Effects of Toxic Divalent Cations on Contractile Proteins with Implications for the Heart: Unraveling Mechanisms of Dysfunction.Int J Mol Sci. 2023 Jun 24;24(13):10579. doi: 10.3390/ijms241310579. Int J Mol Sci. 2023. PMID: 37445756 Free PMC article. Review.
-
The Local Environment of Loop Switch 1 Modulates the Rate of ATP-Induced Dissociation of Human Cardiac Actomyosin.Int J Mol Sci. 2022 Jan 22;23(3):1220. doi: 10.3390/ijms23031220. Int J Mol Sci. 2022. PMID: 35163146 Free PMC article.
-
Switch-1 instability at the active site decouples ATP hydrolysis from force generation in myosin II.Cytoskeleton (Hoboken). 2021 Jan;78(1):3-13. doi: 10.1002/cm.21650. Epub 2021 Jan 11. Cytoskeleton (Hoboken). 2021. PMID: 33381891 Free PMC article.
-
Altered chemomechanical coupling causes impaired motility of the kinesin-4 motors KIF27 and KIF7.J Cell Biol. 2018 Apr 2;217(4):1319-1334. doi: 10.1083/jcb.201708179. Epub 2018 Jan 19. J Cell Biol. 2018. PMID: 29351996 Free PMC article.
References
-
- Marx A., Müller J., Mandelkow E. (2005) The structure of microtubule motor proteins. Adv. Protein Chem. 71, 299–344 - PubMed
-
- Vale R. D., Milligan R. A. (2000) The way things move: looking under the hood of molecular motor proteins. Science 288, 88–95 - PubMed
-
- Sprang S. R. (1997) G protein mechanisms: insights from structural analysis. Annu. Rev. Biochem. 66, 639–678 - PubMed
-
- Kull F. J., Vale R. D., Fletterick R. J. (1998) The case for a common ancestor: kinesin and myosin motor proteins and G proteins. J. Muscle Res. Cell Motil. 19, 877–886 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
