

Caged naloxone reveals opioid signaling deactivation kinetics
- PMID: 23960100
- PMCID: PMC3807075
- DOI: 10.1124/mol.113.088096
Caged naloxone reveals opioid signaling deactivation kinetics
Abstract
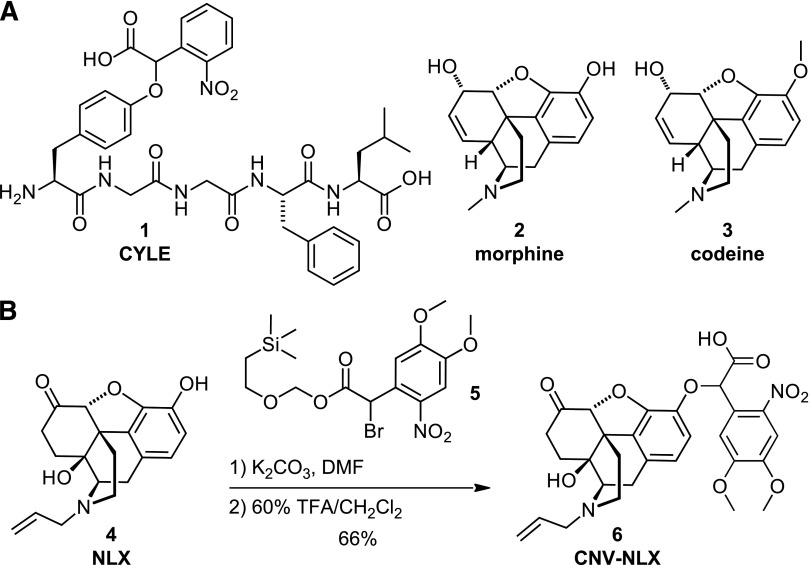
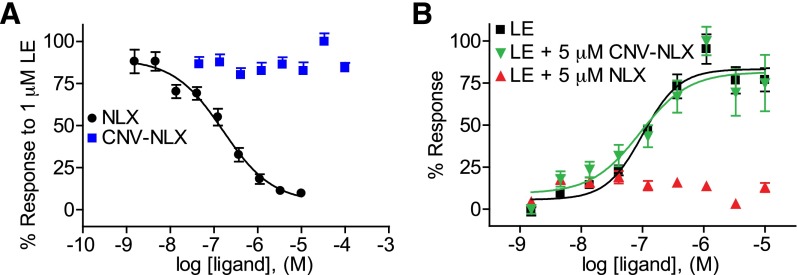
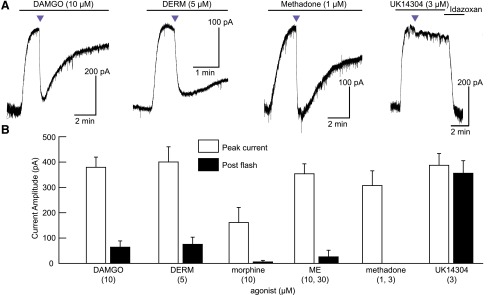
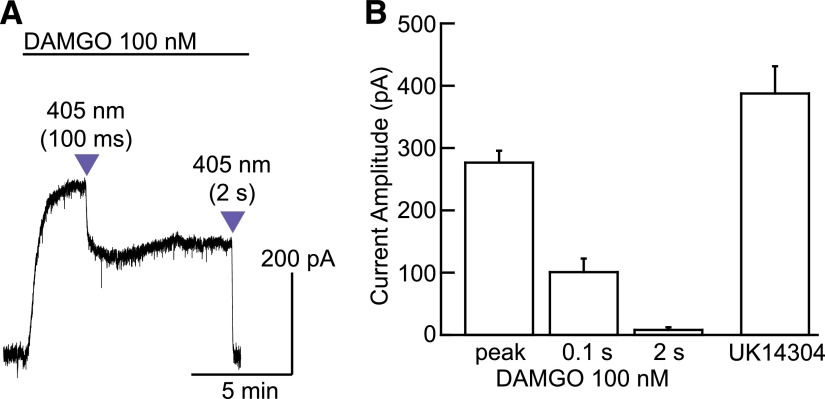
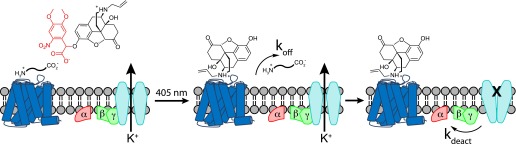
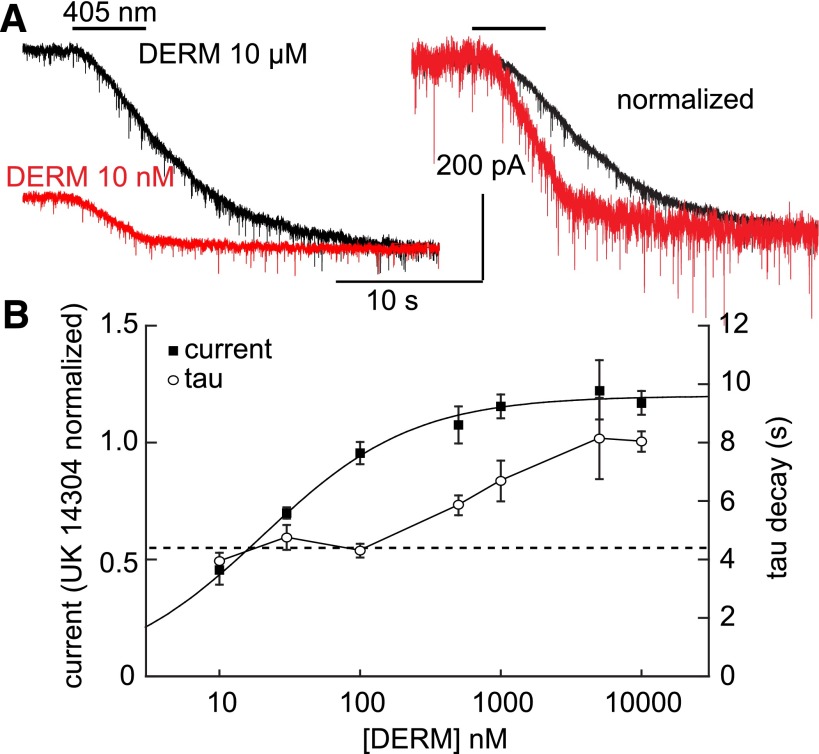
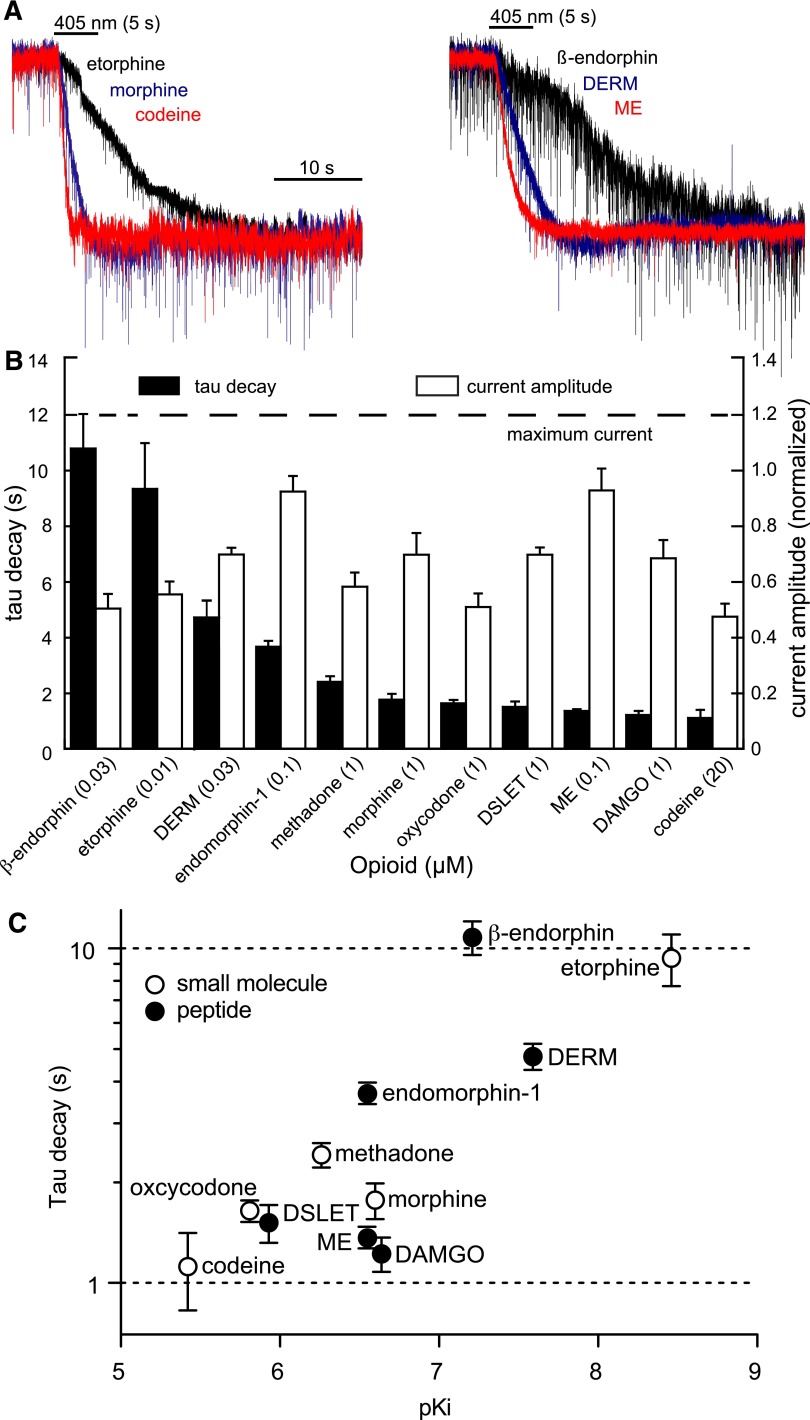
The spatiotemporal dynamics of opioid signaling in the brain remain poorly defined. Photoactivatable opioid ligands provide a means to quantitatively measure these dynamics and their underlying mechanisms in brain tissue. Although activation kinetics can be assessed using caged agonists, deactivation kinetics are obscured by slow clearance of agonist in tissue. To reveal deactivation kinetics of opioid signaling we developed a caged competitive antagonist that can be quickly photoreleased in sufficient concentrations to render agonist dissociation effectively irreversible. Carboxynitroveratryl-naloxone (CNV-NLX), a caged analog of the competitive opioid antagonist NLX, was readily synthesized from commercially available NLX in good yield and found to be devoid of antagonist activity at heterologously expressed opioid receptors. Photolysis in slices of rat locus coeruleus produced a rapid inhibition of the ionic currents evoked by multiple agonists of the μ-opioid receptor (MOR), but not of α-adrenergic receptors, which activate the same pool of ion channels. Using the high-affinity peptide agonist dermorphin, we established conditions under which light-driven deactivation rates are independent of agonist concentration and thus intrinsic to the agonist-receptor complex. Under these conditions, some MOR agonists yielded deactivation rates that are limited by G protein signaling, whereas others appeared limited by agonist dissociation. Therefore, the choice of agonist determines which feature of receptor signaling is unmasked by CNV-NLX photolysis.
Figures
References
-
- Albizu L, Balestre MN, Breton C, Pin JP, Manning M, Mouillac B, Barberis C, Durroux T. (2006) Probing the existence of G protein-coupled receptor dimers by positive and negative ligand-dependent cooperative binding. Mol Pharmacol 70:1783–1791 - PubMed
-
- Childers SR, Snyder SH. (1980) Differential regulation by guanine nucleotides or opiate agonist and antagonist receptor interactions. J Neurochem 34:583–593 - PubMed
-
- Connor M, Traynor J. (2010) Constitutively active μ-opioid receptors. Methods Enzymol 484:445–469 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
