

Acetylation of the KXGS motifs in tau is a critical determinant in modulation of tau aggregation and clearance
- PMID: 23962722
- PMCID: PMC3857946
- DOI: 10.1093/hmg/ddt402
Acetylation of the KXGS motifs in tau is a critical determinant in modulation of tau aggregation and clearance
Abstract
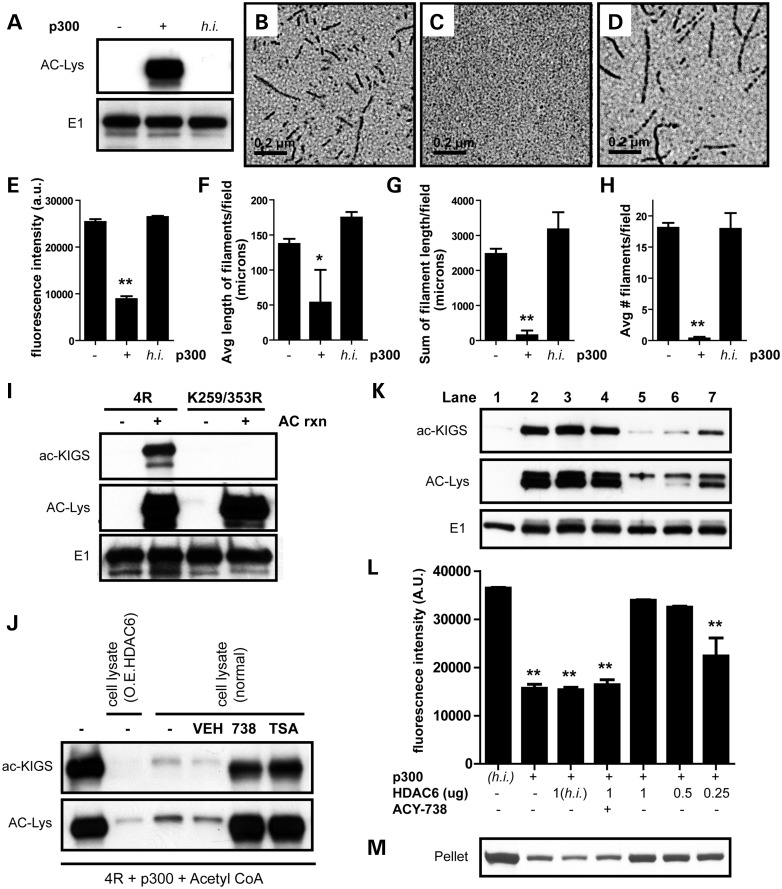
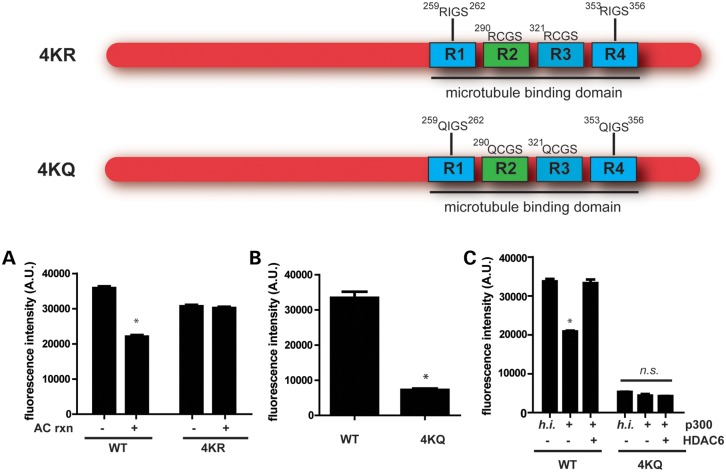
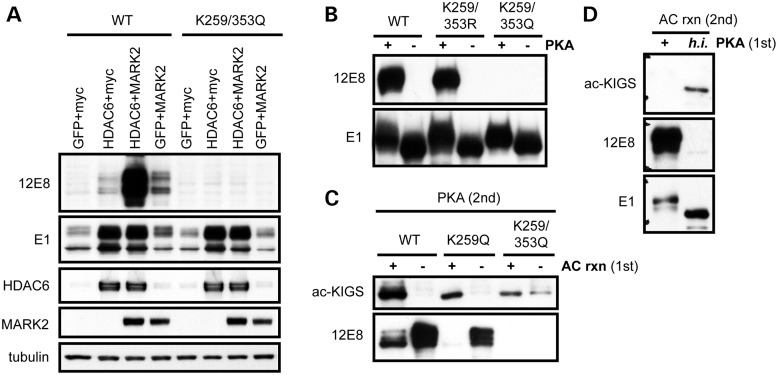
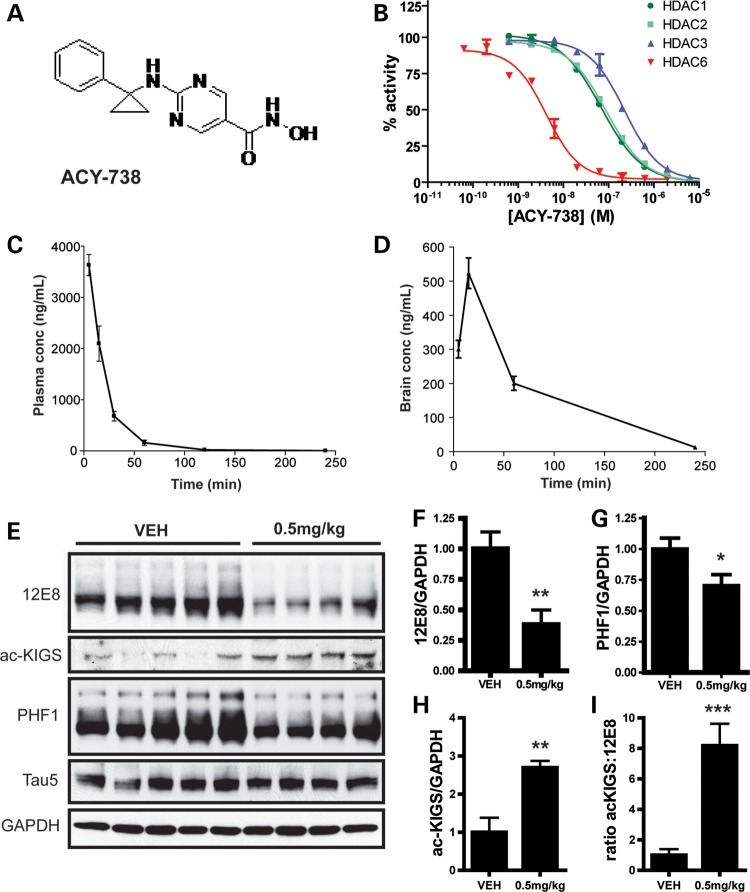
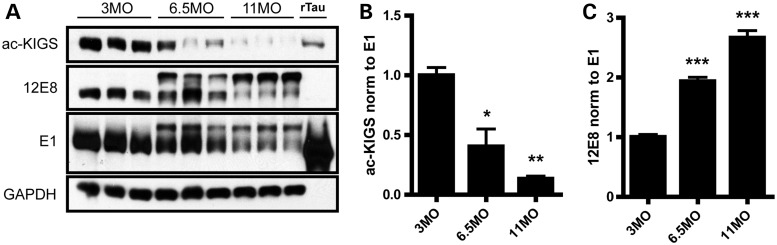
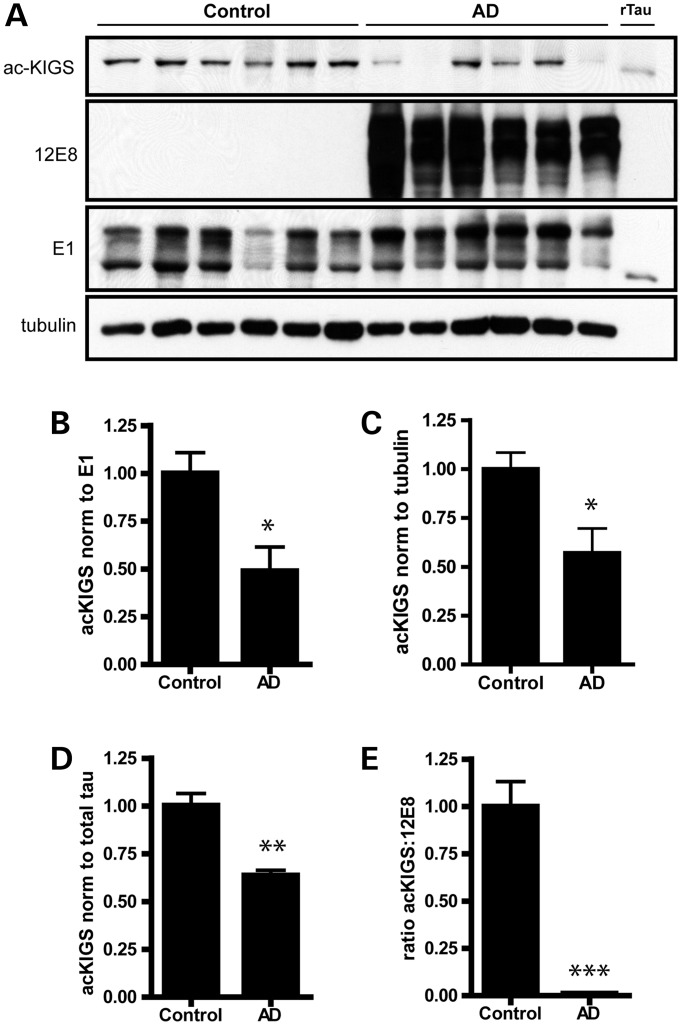
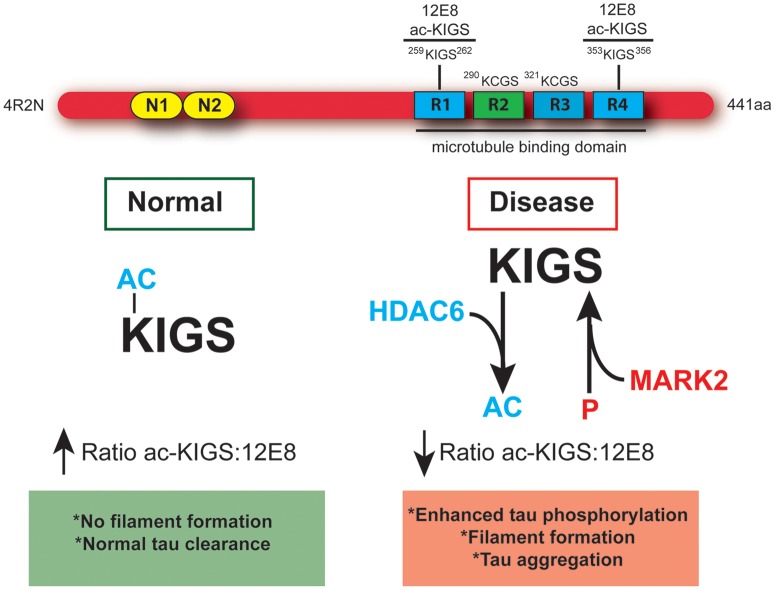
The accumulation of hyperphosphorylated tau in neurofibrillary tangles (NFTs) is a neuropathological hallmark of tauopathies, including Alzheimer's disease (AD) and chronic traumatic encephalopathy, but effective therapies directly targeting the tau protein are currently lacking. Herein, we describe a novel mechanism in which the acetylation of tau on KXGS motifs inhibits phosphorylation on this same motif, and also prevents tau aggregation. Using a site-specific antibody to detect acetylation of KXGS motifs, we demonstrate that these sites are hypoacetylated in patients with AD, as well as a mouse model of tauopathy, suggesting that loss of acetylation on KXGS motifs renders tau vulnerable to pathogenic insults. Furthermore, we identify histone deacetylase 6 (HDAC6) as the enzyme responsible for the deacetylation of these residues, and provide proof of concept that acute treatment with a selective and blood-brain barrier-permeable HDAC6 inhibitor enhances acetylation and decreases phosphorylation on tau's KXGS motifs in vivo. As such, we have uncovered a novel therapeutic pathway that can be manipulated to block the formation of pathogenic tau species in disease.
Figures
References
-
- Buee L., Bussiere T., Buee-Scherrer V., Delacourte A., Hof P.R. Tau protein isoforms, phosphorylation and role in neurodegenerative disorders. Brain Res. Brain Res. Rev. 2000;33:95–130. doi:10.1016/S0165-0173(00)00019-9. - DOI - PubMed
-
- Dickson D.W. Tau and synuclein and their role in neuropathology. Brain Pathol. 1999;9:657–661. doi:10.1111/j.1750-3639.1999.tb00548.x. - DOI - PMC - PubMed
-
- McKee A.C., Cantu R.C., Nowinski C.J., Hedley-Whyte E.T., Gavett B.E., Budson A.E., Santini V.E., Lee H.S., Kubilus C.A., Stern R.A. Chronic traumatic encephalopathy in athletes: progressive tauopathy after repetitive head injury. J. Neuropathol. Exp. Neurol. 2009;68:709–735. doi:10.1097/NEN.0b013e3181a9d503. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
