

Mediator is an intrinsic component of the basal RNA polymerase II machinery in vivo
- PMID: 23963697
- PMCID: PMC3834807
- DOI: 10.1093/nar/gkt701
Mediator is an intrinsic component of the basal RNA polymerase II machinery in vivo
Abstract
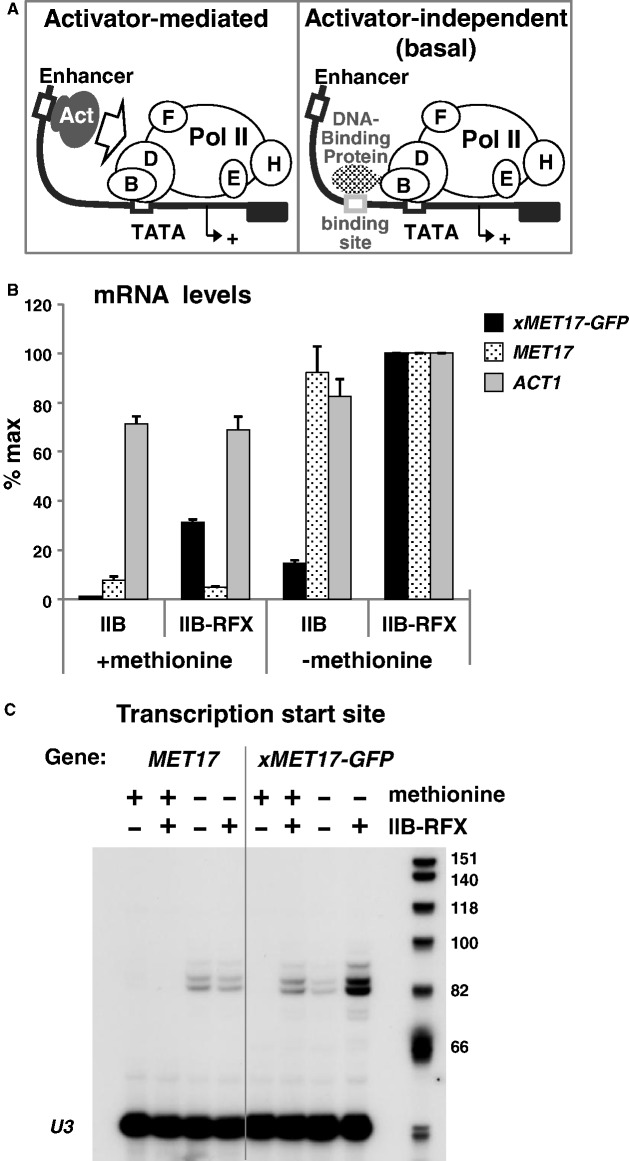
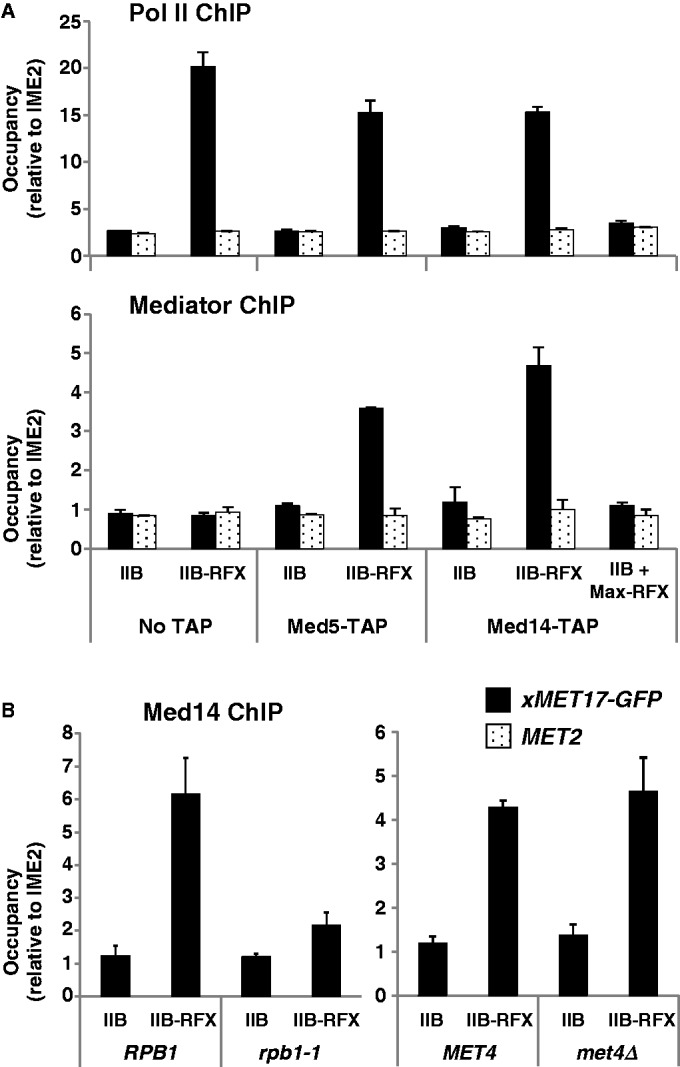
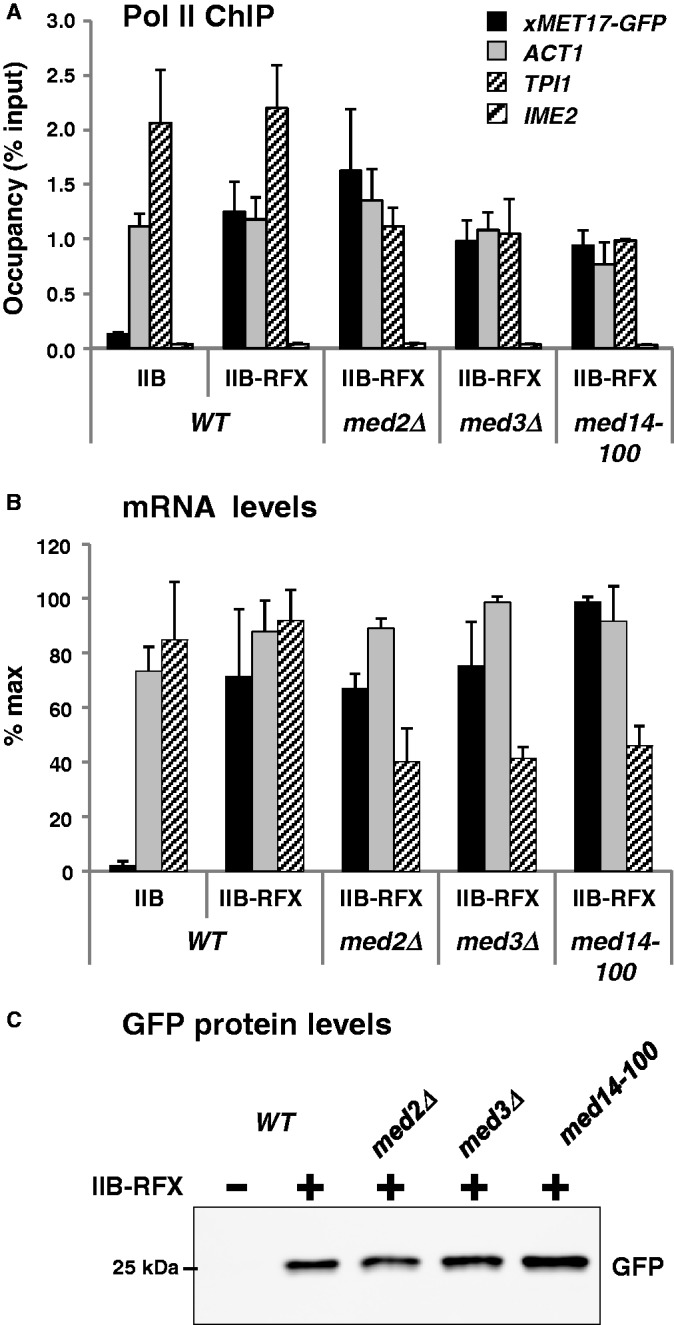
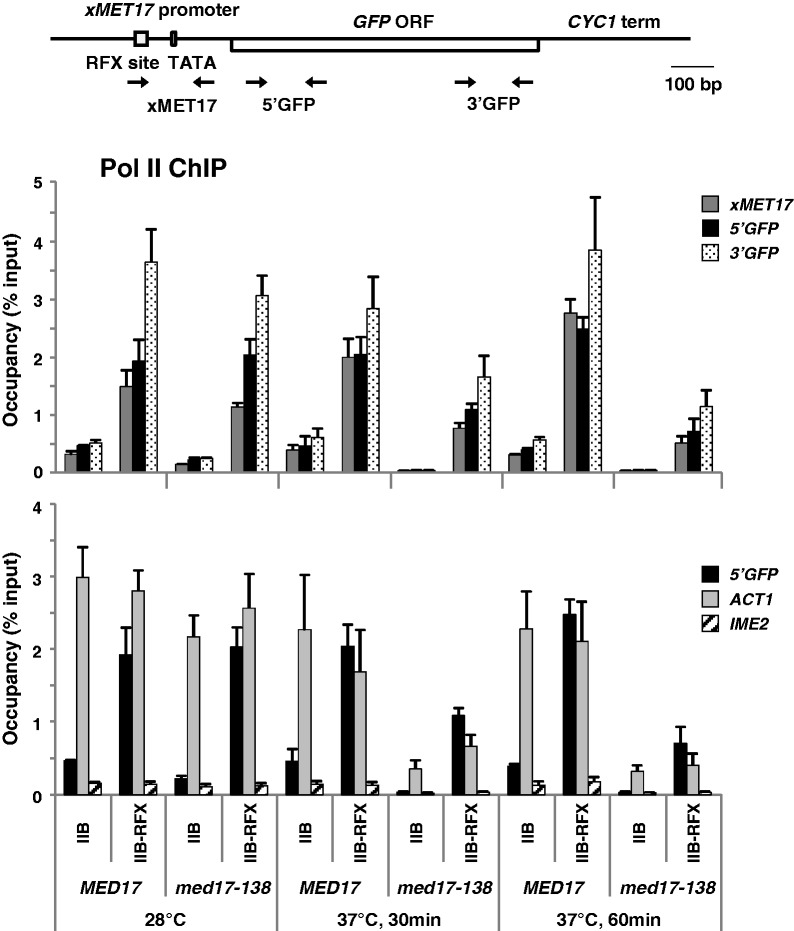
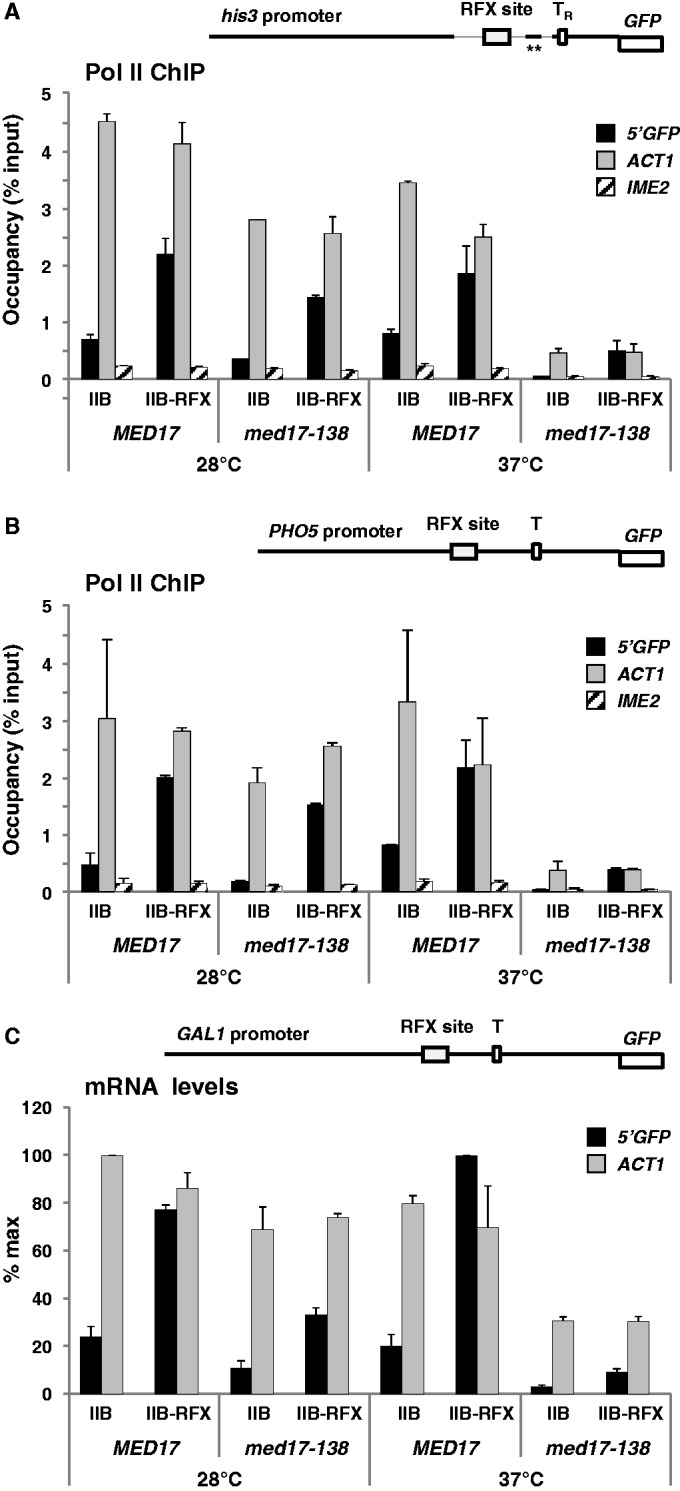
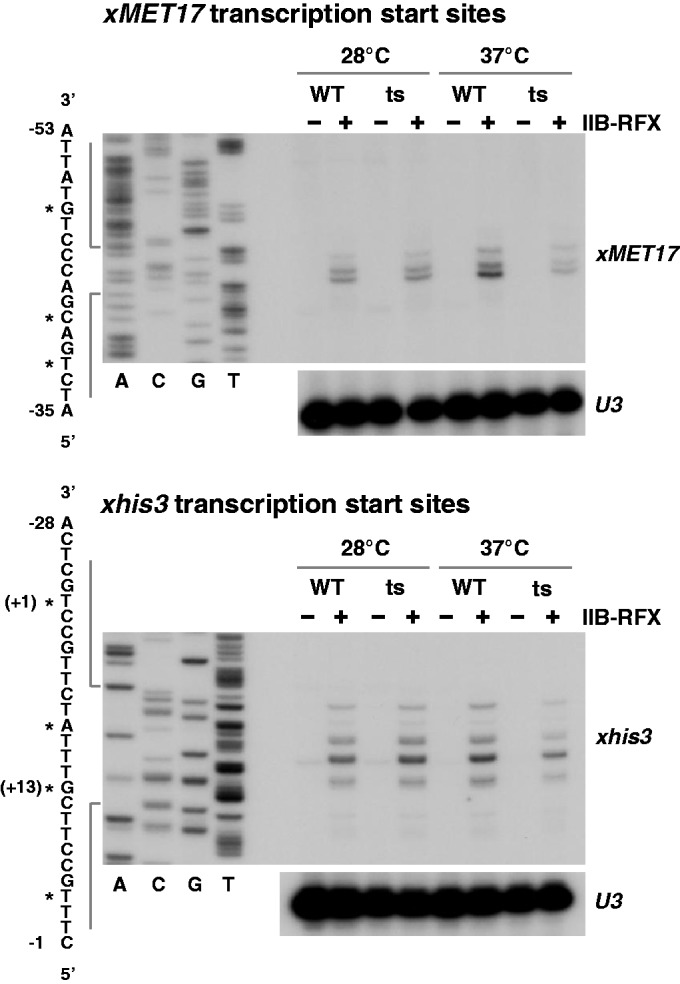
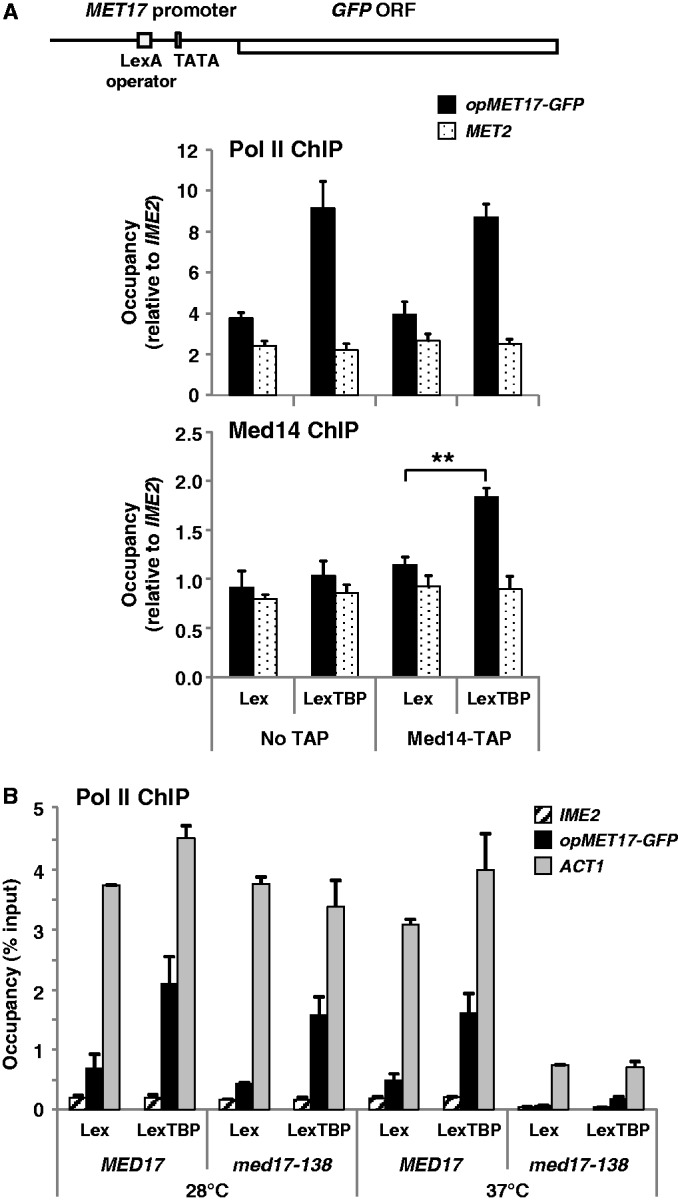
Mediator is a prominent multisubunit coactivator that functions as a bridge between gene-specific activators and the basal RNA polymerase (Pol) II initiation machinery. Here, we study the poorly documented role of Mediator in basal, or activator-independent, transcription in vivo. We show that Mediator is still present at the promoter when the Pol II machinery is recruited in the absence of an activator, in this case through a direct fusion between a basal transcription factor and a heterologous DNA binding protein bound to the promoter. Moreover, transcription resulting from activator-independent recruitment of the Pol II machinery is impaired by inactivation of the essential Mediator subunit Med17 due to the loss of Pol II from the promoter. Our results strongly support that Mediator is an integral component of the minimal machinery essential in vivo for stable Pol II association with the promoter.
Figures
References
-
- Kornberg RD. Mediator and the mechanism of transcriptional activation. Trends Biochem. Sci. 2005;30:235–239. - PubMed
-
- Davis JA, Takagi Y, Kornberg RD, Asturias FA. Structure of the yeast RNA polymerase II holoenzyme: mediator conformation and polymerase interaction. Mol. Cell. 2002;10:409–415. - PubMed
-
- Borggrefe T, Davis R, Erdjument-Bromage H, Tempst P, Kornberg RD. A complex of the Srb8, -9, -10, and -11 transcriptional regulatory proteins from yeast. J. Biol. Chem. 2002;277:44202–44207. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
