

fMRI orientation decoding in V1 does not require global maps or globally coherent orientation stimuli
- PMID: 23964251
- PMCID: PMC3740242
- DOI: 10.3389/fpsyg.2013.00493
fMRI orientation decoding in V1 does not require global maps or globally coherent orientation stimuli
Abstract
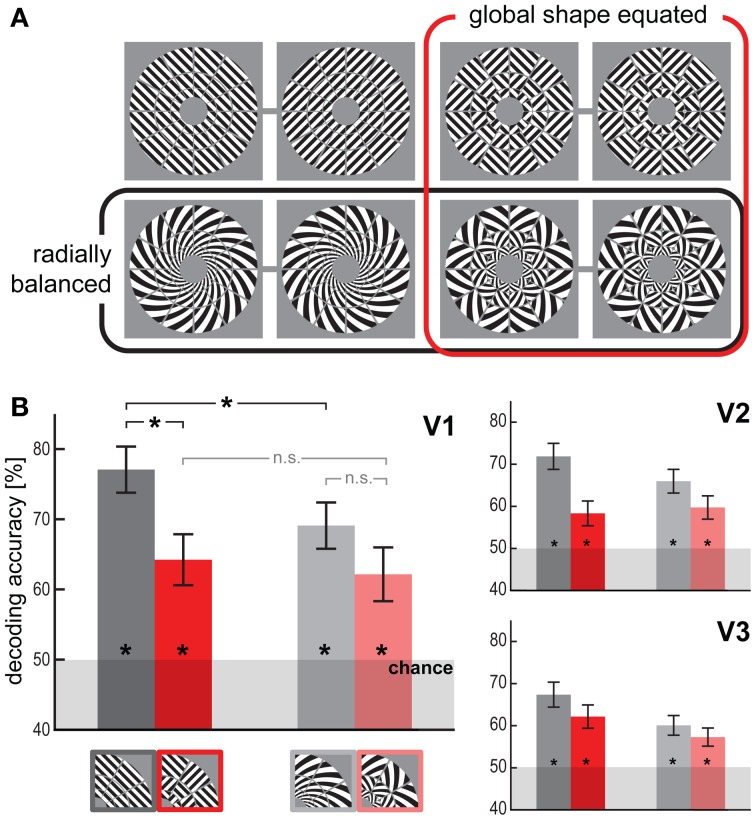
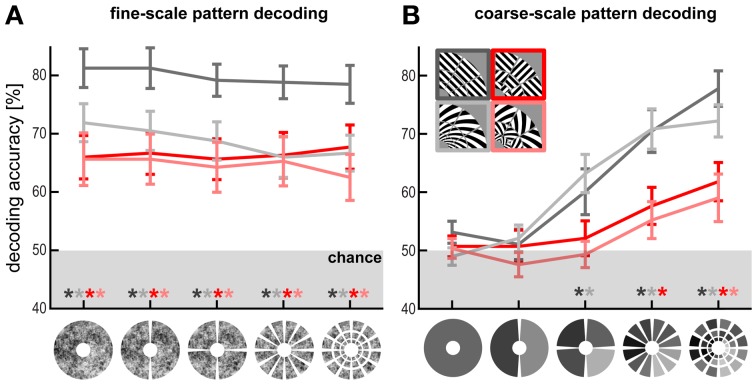
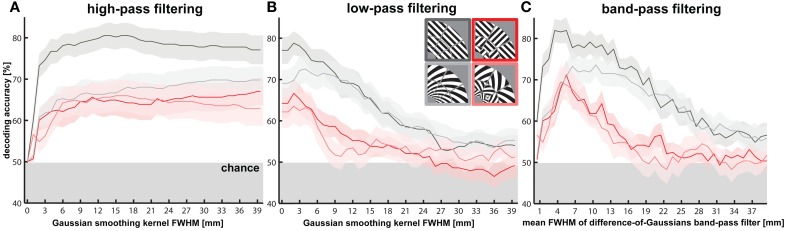
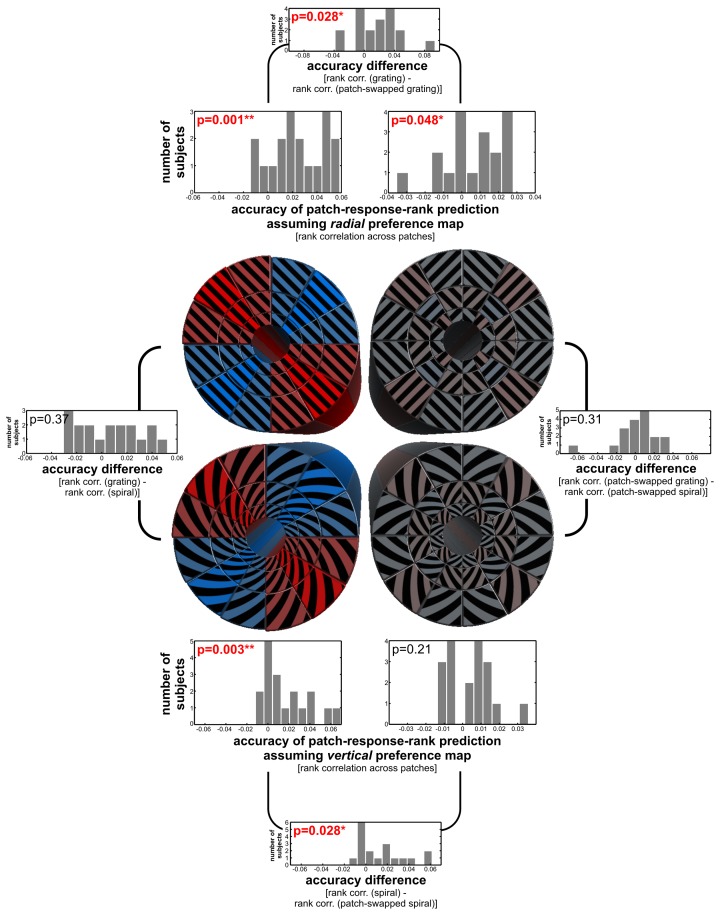
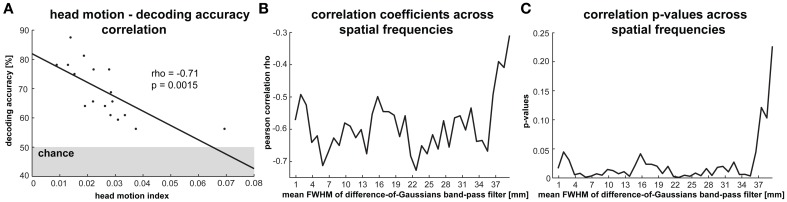
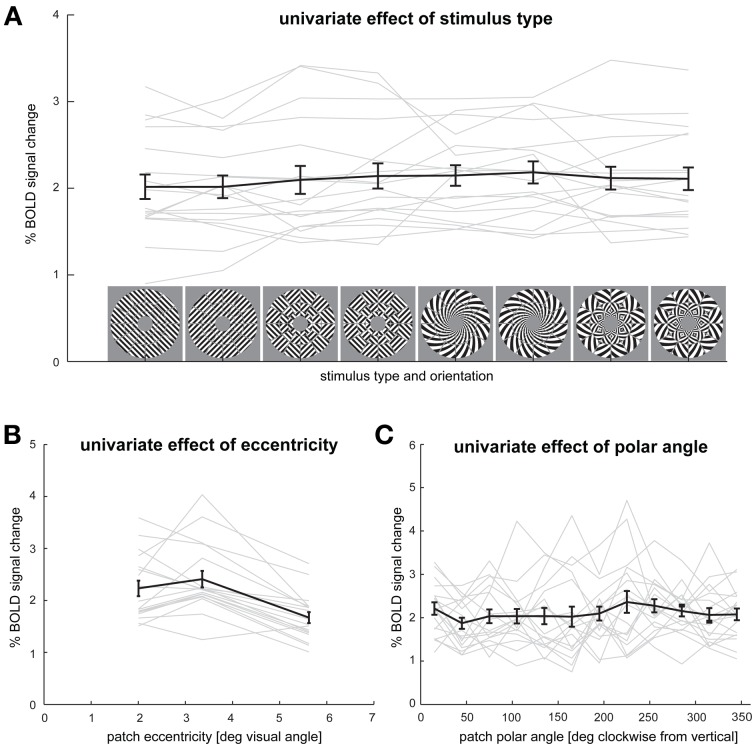
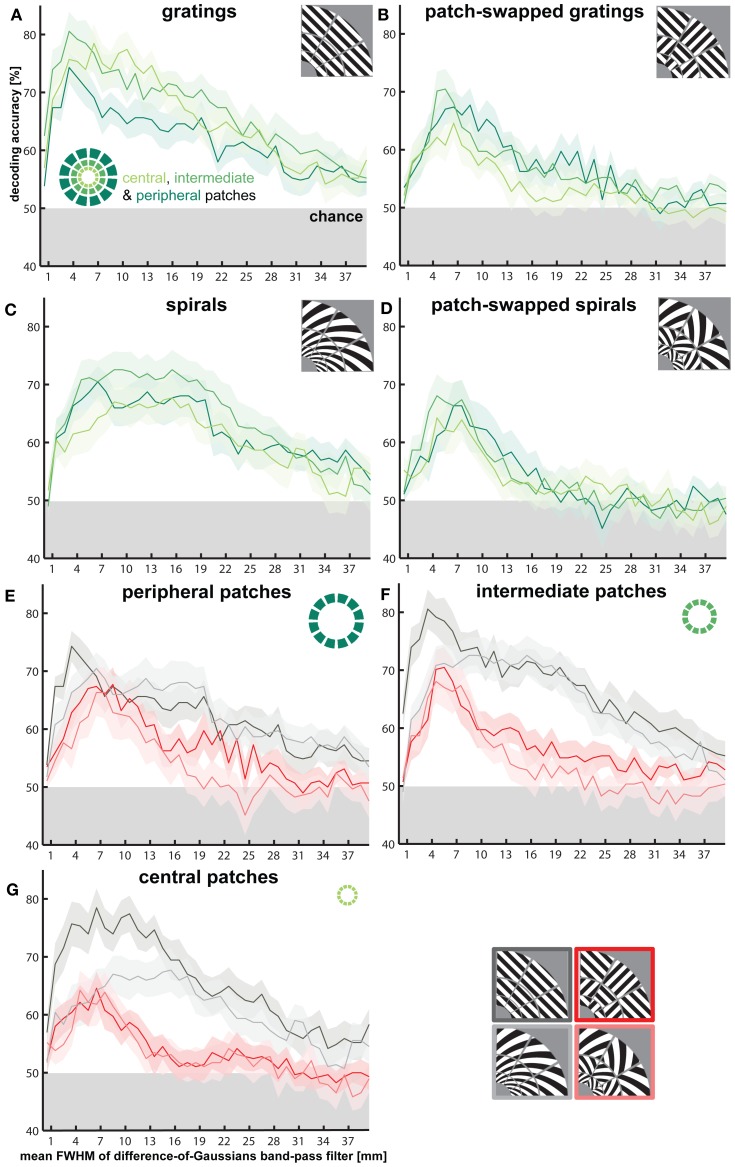
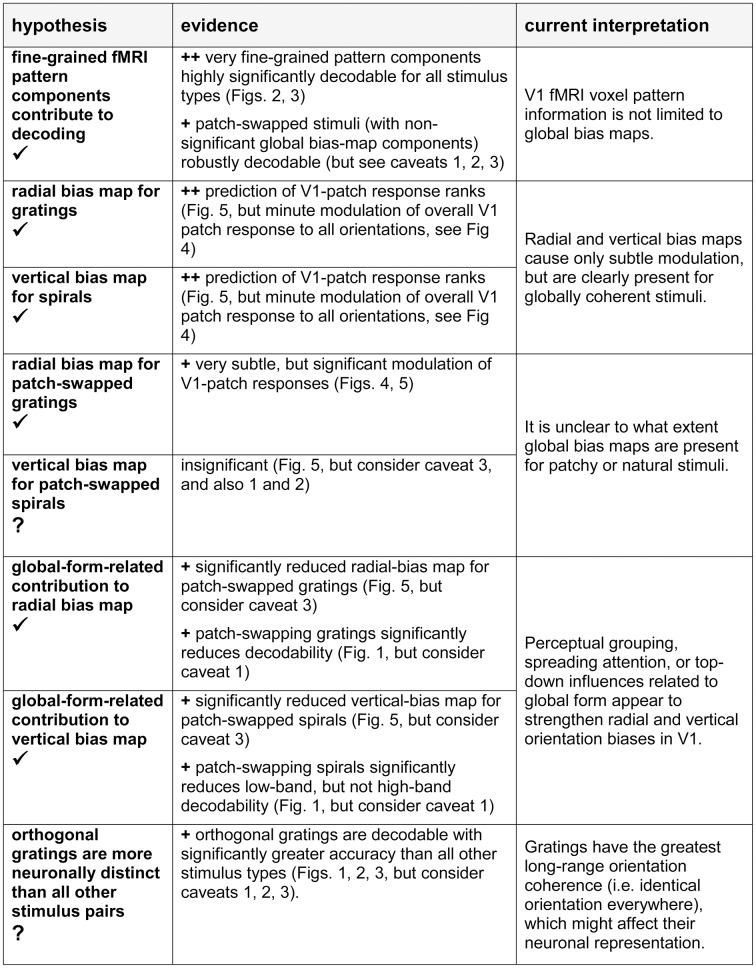
The orientation of a large grating can be decoded from V1 functional magnetic resonance imaging (fMRI) data, even at low resolution (3-mm isotropic voxels). This finding has suggested that columnar-level neuronal information might be accessible to fMRI at 3T. However, orientation decodability might alternatively arise from global orientation-preference maps. Such global maps across V1 could result from bottom-up processing, if the preferences of V1 neurons were biased toward particular orientations (e.g., radial from fixation, or cardinal, i.e., vertical or horizontal). Global maps could also arise from local recurrent or top-down processing, reflecting pre-attentive perceptual grouping, attention spreading, or predictive coding of global form. Here we investigate whether fMRI orientation decoding with 2-mm voxels requires (a) globally coherent orientation stimuli and/or (b) global-scale patterns of V1 activity. We used opposite-orientation gratings (balanced about the cardinal orientations) and spirals (balanced about the radial orientation), along with novel patch-swapped variants of these stimuli. The two stimuli of a patch-swapped pair have opposite orientations everywhere (like their globally coherent parent stimuli). However, the two stimuli appear globally similar, a patchwork of opposite orientations. We find that all stimulus pairs are robustly decodable, demonstrating that fMRI orientation decoding does not require globally coherent orientation stimuli. Furthermore, decoding remained robust after spatial high-pass filtering for all stimuli, showing that fine-grained components of the fMRI patterns reflect visual orientations. Consistent with previous studies, we found evidence for global radial and vertical preference maps in V1. However, these were weak or absent for patch-swapped stimuli, suggesting that global preference maps depend on globally coherent orientations and might arise through recurrent or top-down processes related to the perception of global form.
Keywords: decoding; fMRI; global form; hyperacuity; orientation selectivity; pattern analysis; radial bias; visual cortex.
Figures

References
-
- Chang C. C., Lin C. J. (2011). LIBSVM: A Library for Support Vector Machines. ACM Transactions on Intelligent Systems and Technology (TIST). (New York, NY: Thieme Medical Publishers; ), 10.1145/1961189.1961199 - DOI
-
- Clifford C. W. G., Mannion D., Seymour K., McDonald J., Bartels A. (2011). Are Coarse-Scale Orientation Maps Really Necessary for Orientation Decoding? Available online at: http://www.jneurosci.org/content/31/13/4792/reply
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources