

Wall teichoic acid structure governs horizontal gene transfer between major bacterial pathogens
- PMID: 23965785
- PMCID: PMC3903184
- DOI: 10.1038/ncomms3345
Wall teichoic acid structure governs horizontal gene transfer between major bacterial pathogens
Abstract
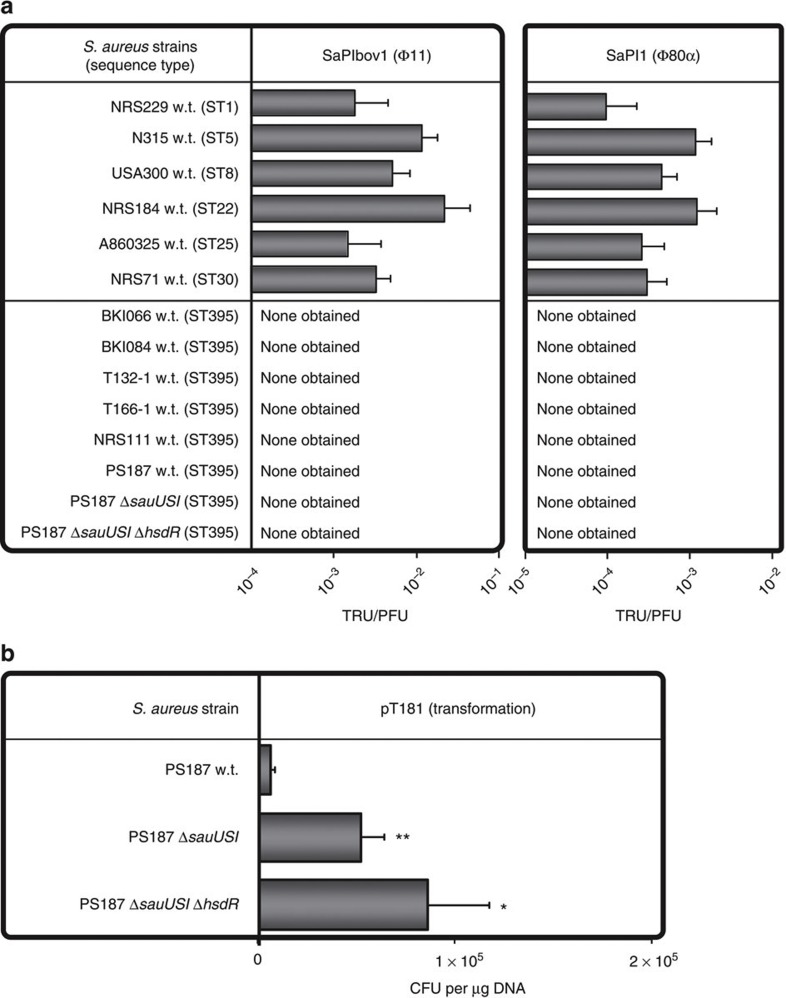
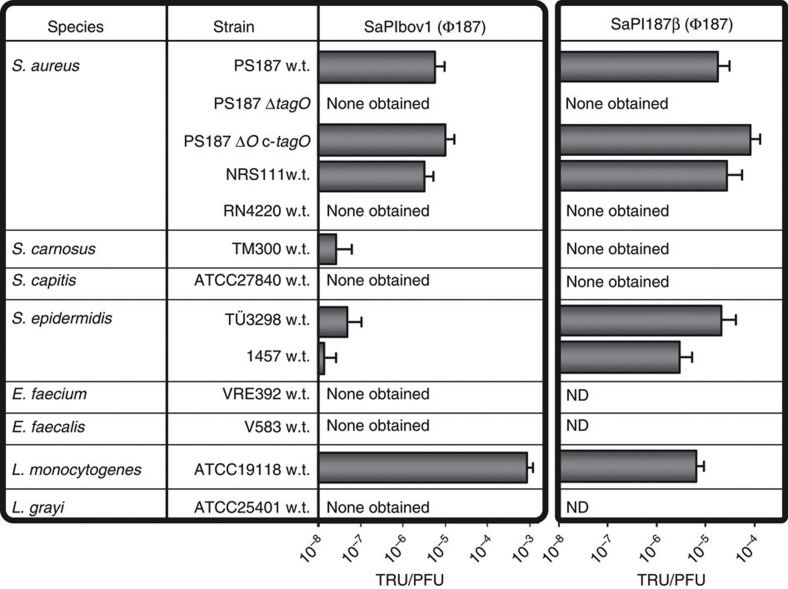
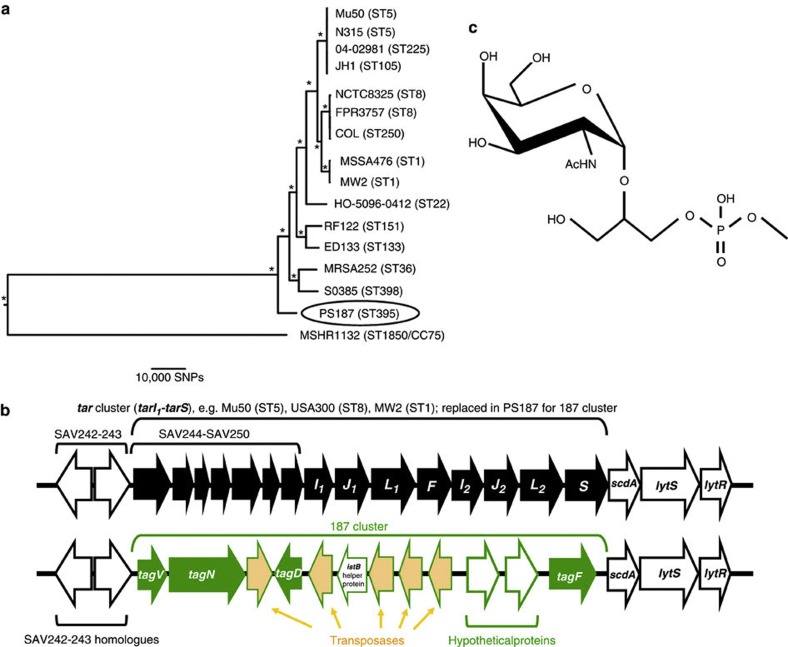
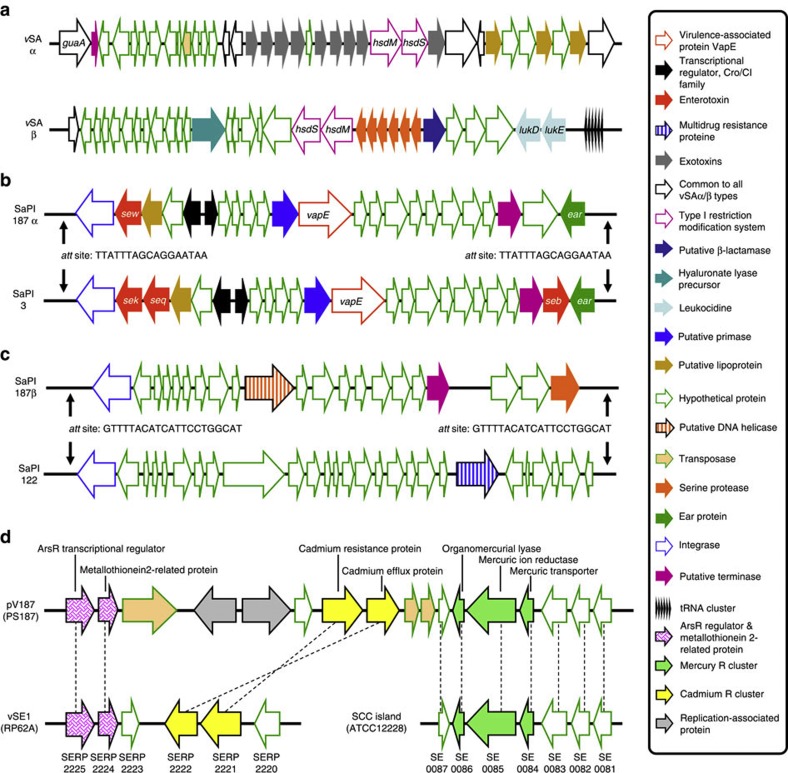
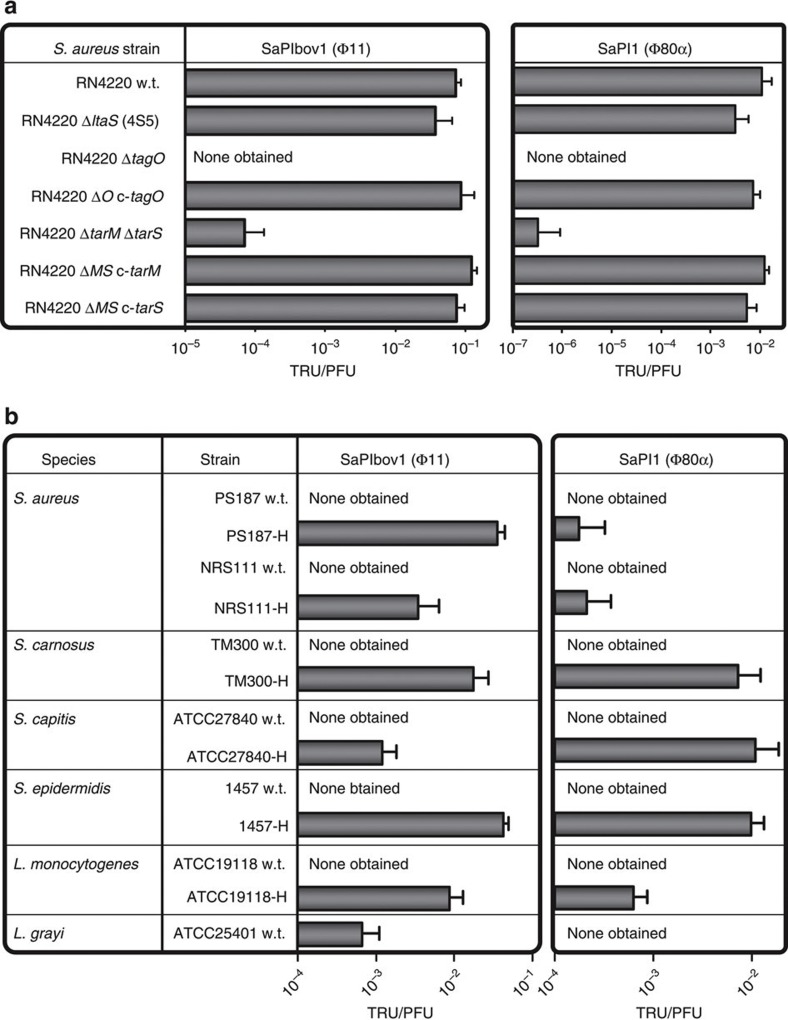
Mobile genetic elements (MGEs) encoding virulence and resistance genes are widespread in bacterial pathogens, but it has remained unclear how they occasionally jump to new host species. Staphylococcus aureus clones exchange MGEs such as S. aureus pathogenicity islands (SaPIs) with high frequency via helper phages. Here we report that the S. aureus ST395 lineage is refractory to horizontal gene transfer (HGT) with typical S. aureus but exchanges SaPIs with other species and genera including Staphylococcus epidermidis and Listeria monocytogenes. ST395 produces an unusual wall teichoic acid (WTA) resembling that of its HGT partner species. Notably, distantly related bacterial species and genera undergo efficient HGT with typical S. aureus upon ectopic expression of S. aureus WTA. Combined with genomic analyses, these results indicate that a 'glycocode' of WTA structures and WTA-binding helper phages permits HGT even across long phylogenetic distances thereby shaping the evolution of Gram-positive pathogens.
Figures
References
-
- Thomas C. M. & Nielsen K. M. Mechanisms of, and barriers to, horizontal gene transfer between bacteria. Nat. Rev. Microbiol. 3, 711–721 (2005). - PubMed
-
- Frost L. S., Leplae R., Summers A. O. & Toussaint A. Mobile genetic elements: the agents of open source evolution. Nat. Rev. Microbiol. 3, 722–732 (2005). - PubMed
-
- Lindsay J. A. Genomic variation and evolution of Staphylococcus aureus. Int. J. Med. Microbiol. 300, 98–103 (2010). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
