

Global RNA structure analysis of poliovirus identifies a conserved RNA structure involved in viral replication and infectivity
- PMID: 23966409
- PMCID: PMC3807356
- DOI: 10.1128/JVI.01560-13
Global RNA structure analysis of poliovirus identifies a conserved RNA structure involved in viral replication and infectivity
Abstract
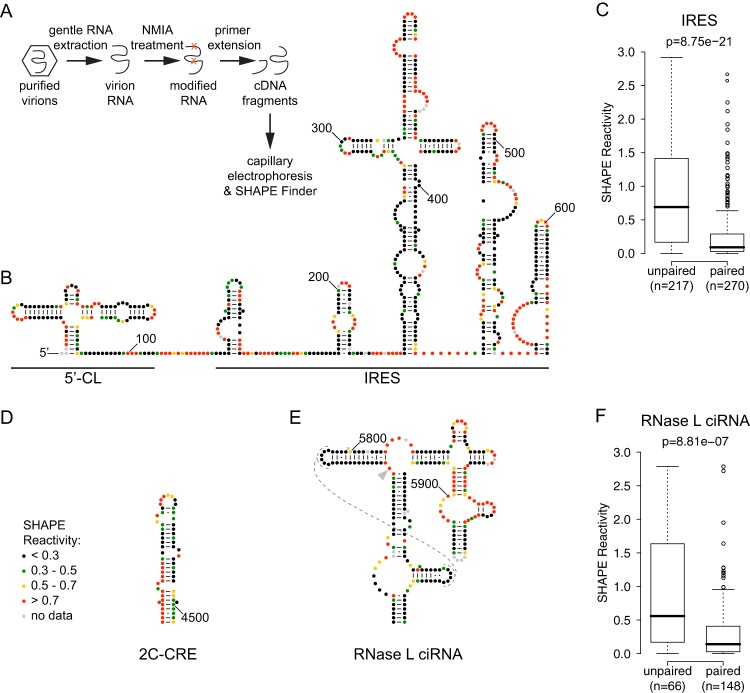
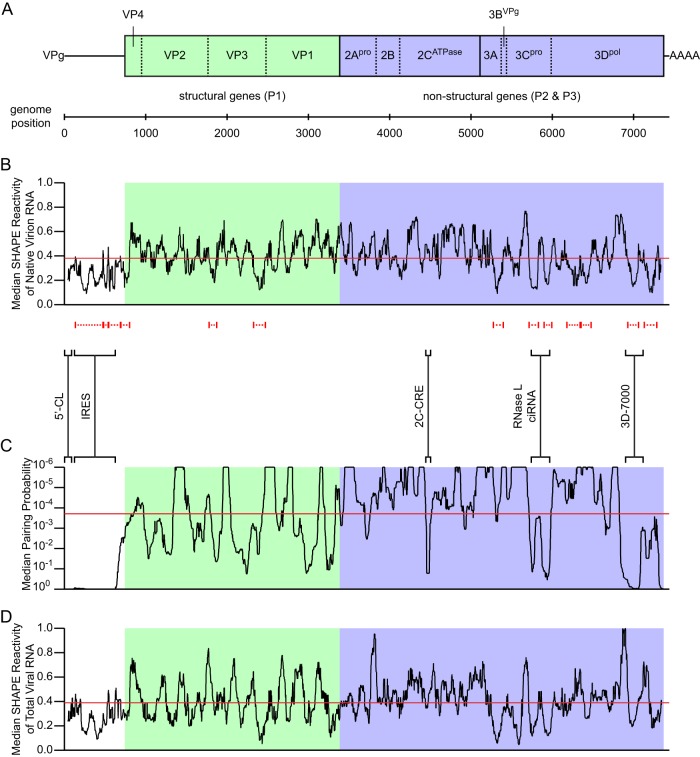
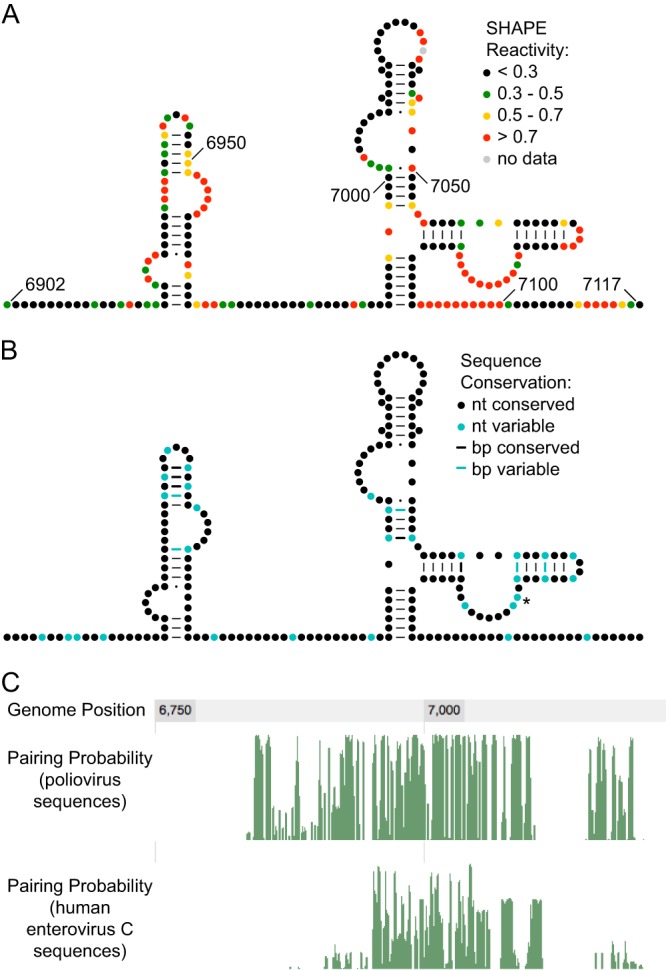
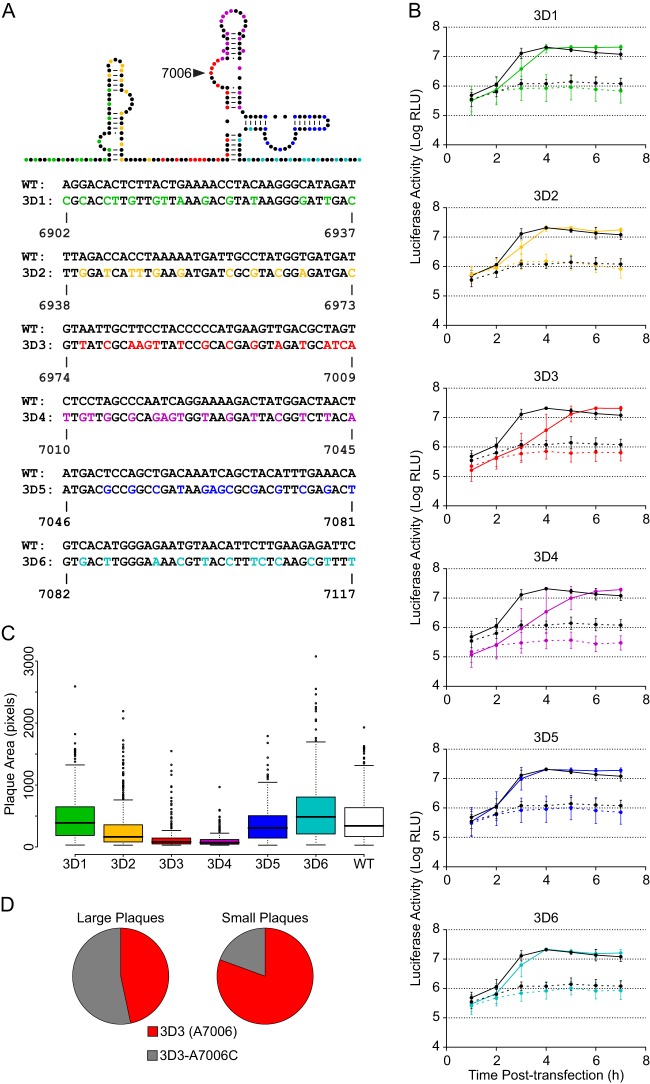
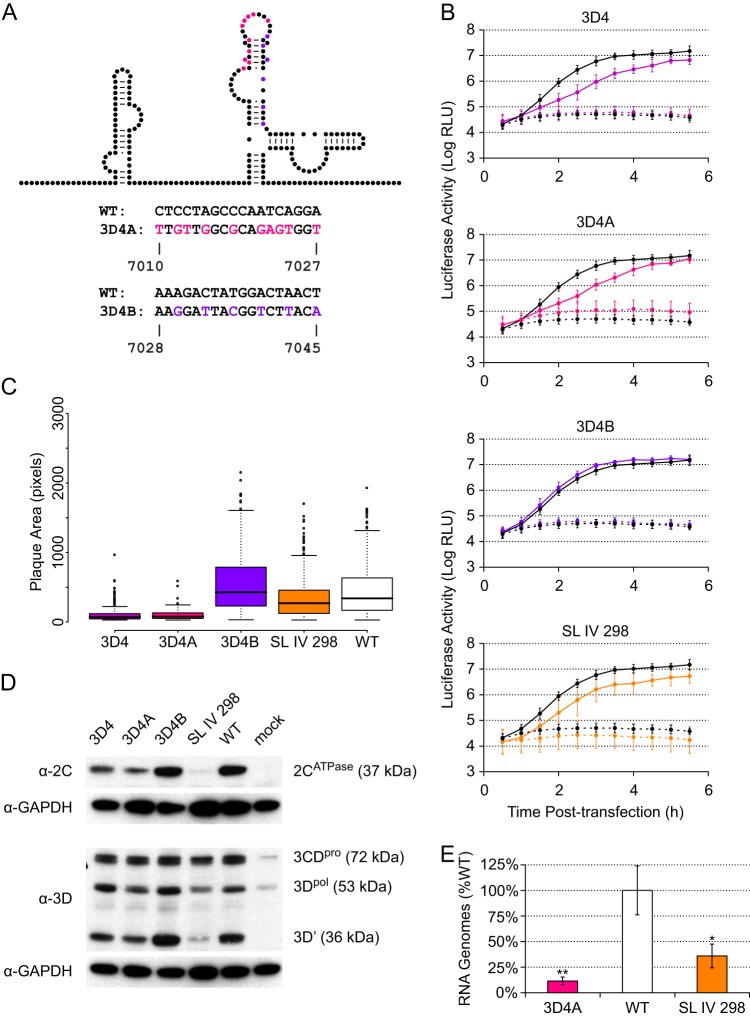
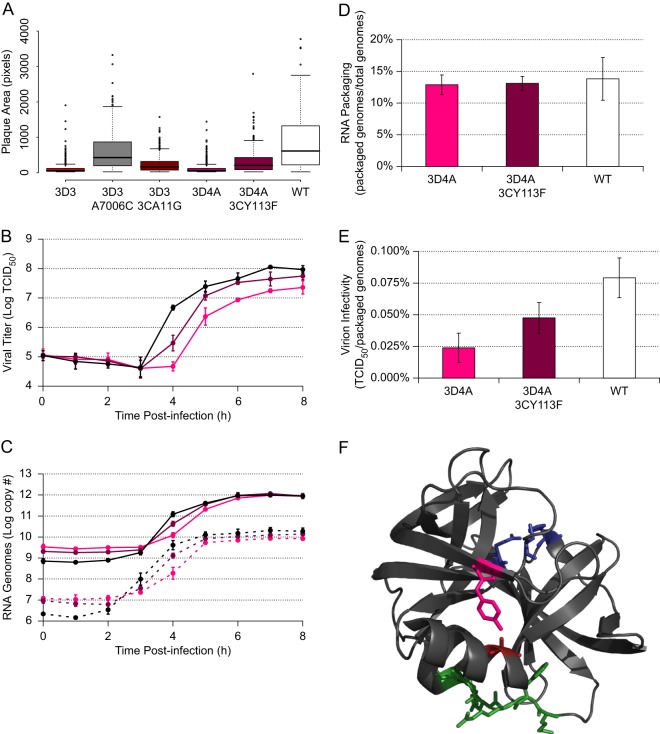
The genomes of RNA viruses often contain RNA structures that are crucial for translation and RNA replication and may play additional, uncharacterized roles during the viral replication cycle. For the picornavirus family member poliovirus, a number of functional RNA structures have been identified, but much of its genome, especially the open reading frame, has remained uncharacterized. We have now generated a global RNA structure map of the poliovirus genome using a chemical probing approach that interrogates RNA structure with single-nucleotide resolution. In combination with orthogonal evolutionary analyses, we uncover several conserved RNA structures in the open reading frame of the viral genome. To validate the ability of our global analyses to identify functionally important RNA structures, we further characterized one of the newly identified structures, located in the region encoding the RNA-dependent RNA polymerase, 3D(pol), by site-directed mutagenesis. Our results reveal that the structure is required for viral replication and infectivity, since synonymous mutants are defective in these processes. Furthermore, these defects can be partially suppressed by mutations in the viral protein 3C(pro), which suggests the existence of a novel functional interaction between an RNA structure in the 3D(pol)-coding region and the viral protein(s) 3C(pro) and/or its precursor 3CD(pro).
Figures
References
-
- Hogle JM, Chow M, Filman DJ. 1985. Three-dimensional structure of poliovirus at 2.9 A resolution. Science 229:1358–1365 - PubMed
-
- Kew OM, Sutter RW, de Gourville EM, Dowdle WR, Pallansch MA. 2005. Vaccine-derived polioviruses and the endgame strategy for global polio eradication. Annu. Rev. Microbiol. 59:587–635 - PubMed
-
- Trono D, Pelletier J, Sonenberg N, Baltimore D. 1988. Translation in mammalian cells of a gene linked to the poliovirus 5′ noncoding region. Science 241:445–448 - PubMed
-
- Andino R, Rieckhof GE, Baltimore D. 1990. A functional ribonucleoprotein complex forms around the 5′ end of poliovirus RNA. Cell 63:369–380 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
