

A Bacteriophage tailspike domain promotes self-cleavage of a human membrane-bound transcription factor, the myelin regulatory factor MYRF
- PMID: 23966832
- PMCID: PMC3742443
- DOI: 10.1371/journal.pbio.1001624
A Bacteriophage tailspike domain promotes self-cleavage of a human membrane-bound transcription factor, the myelin regulatory factor MYRF
Abstract
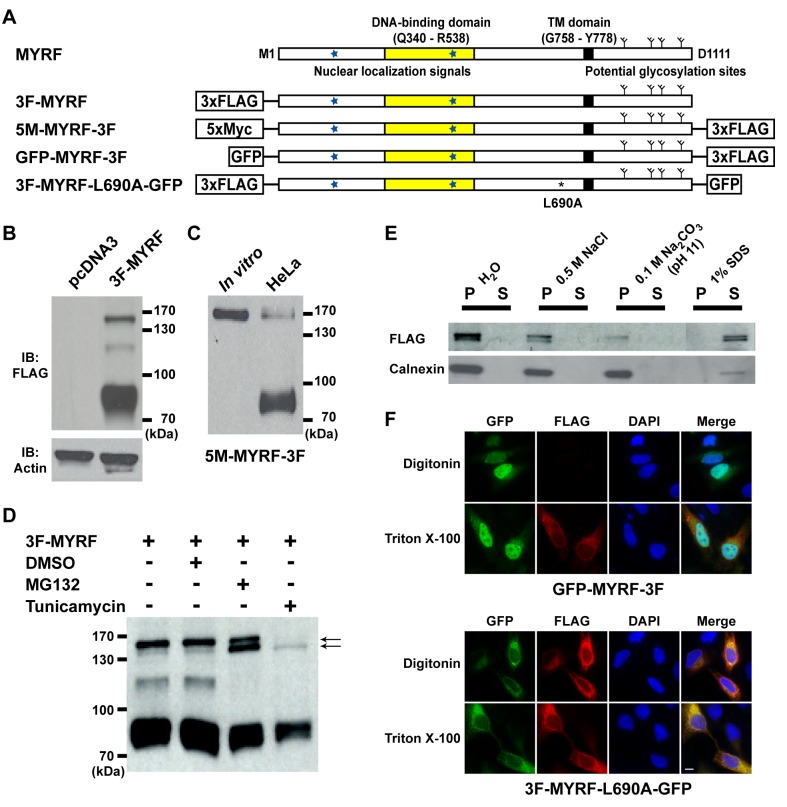
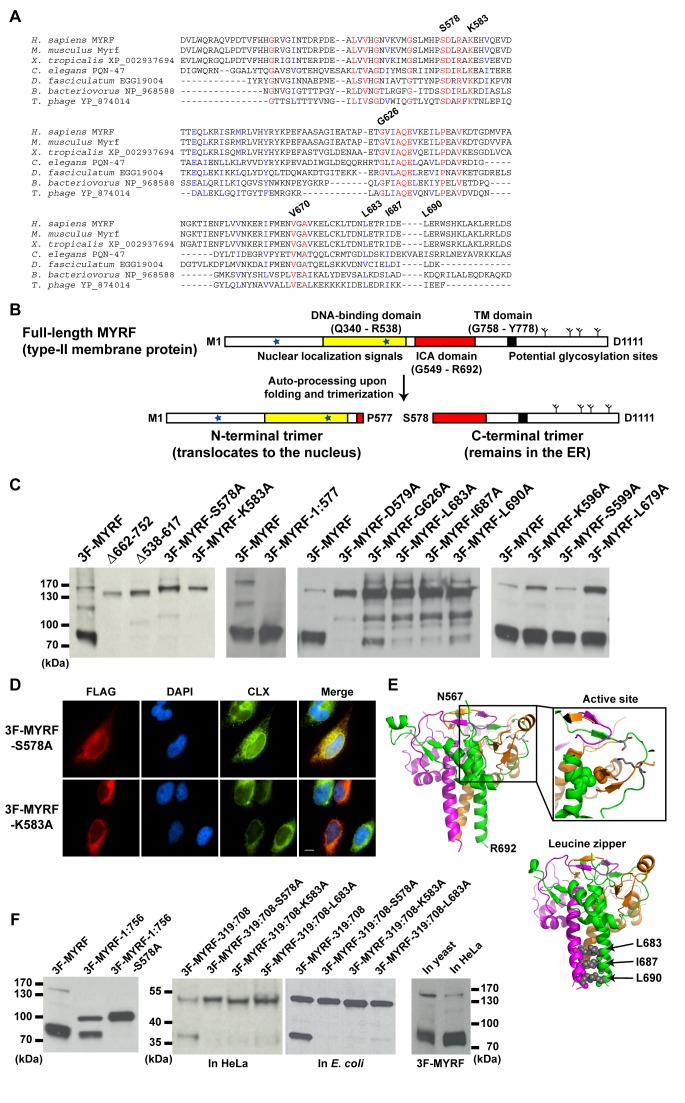
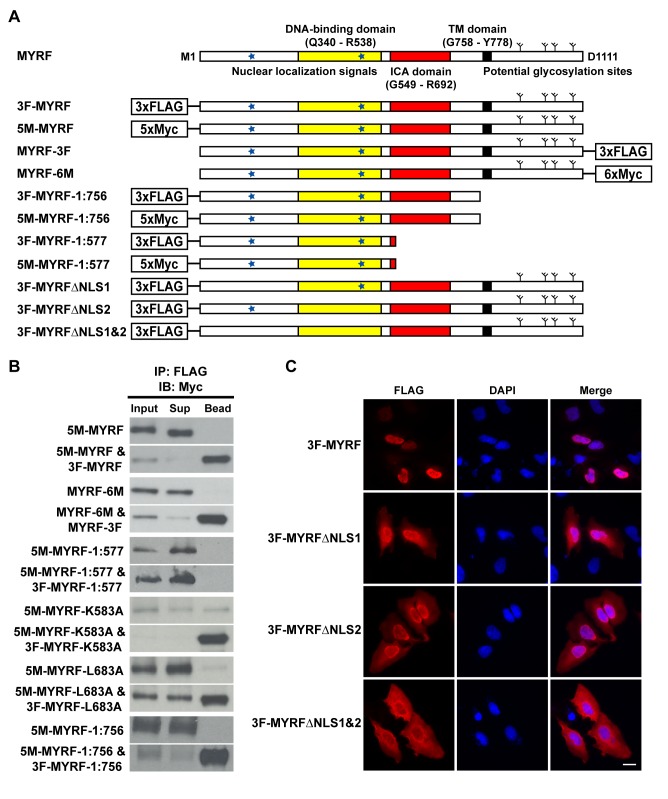
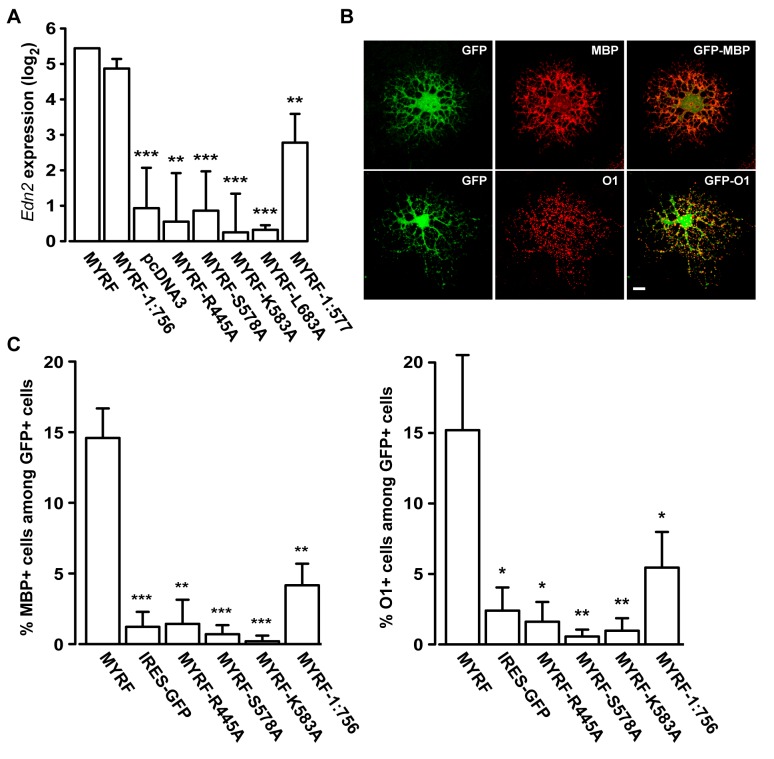
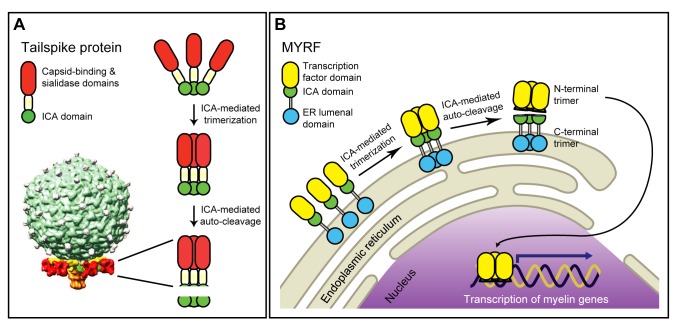
Myelination of the central nervous system (CNS) is critical to vertebrate nervous systems for efficient neural signaling. CNS myelination occurs as oligodendrocytes terminally differentiate, a process regulated in part by the myelin regulatory factor, MYRF. Using bioinformatics and extensive biochemical and functional assays, we find that MYRF is generated as an integral membrane protein that must be processed to release its transcription factor domain from the membrane. In contrast to most membrane-bound transcription factors, MYRF proteolysis seems constitutive and independent of cell- and tissue-type, as we demonstrate by reconstitution in E. coli and yeast. The apparent absence of physiological cues raises the question as to how and why MYRF is processed. By using computational methods capable of recognizing extremely divergent sequence homology, we identified a MYRF protein domain distantly related to bacteriophage tailspike proteins. Although occurring in otherwise unrelated proteins, the phage domains are known to chaperone the tailspike proteins' trimerization and auto-cleavage, raising the hypothesis that the MYRF domain might contribute to a novel activation method for a membrane-bound transcription factor. We find that the MYRF domain indeed serves as an intramolecular chaperone that facilitates MYRF trimerization and proteolysis. Functional assays confirm that the chaperone domain-mediated auto-proteolysis is essential both for MYRF's transcriptional activity and its ability to promote oligodendrocyte maturation. This work thus reveals a previously unknown key step in CNS myelination. These data also reconcile conflicting observations of this protein family, different members of which have been identified as transmembrane or nuclear proteins. Finally, our data illustrate a remarkable evolutionary repurposing between bacteriophages and eukaryotes, with a chaperone domain capable of catalyzing trimerization-dependent auto-proteolysis in two entirely distinct protein and cellular contexts, in one case participating in bacteriophage tailspike maturation and in the other activating a key transcription factor for CNS myelination.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

Comment in
-
Myelination borrows a trick from phage.PLoS Biol. 2013;11(8):e1001626. doi: 10.1371/journal.pbio.1001626. Epub 2013 Aug 13. PLoS Biol. 2013. PMID: 23966834 Free PMC article. No abstract available.
References
-
- Hoppe T, Matuschewski K, Rape M, Schlenker S, Ulrich HD, et al. (2000) Activation of a membrane-bound transcription factor by regulated ubiquitin/proteasome-dependent processing. Cell 102: 577–586. - PubMed
-
- Wang X, Sato R, Brown MS, Hua X, Goldstein JL (1994) SREBP-1, a membrane-bound transcription factor released by sterol-regulated proteolysis. Cell 77: 53–62. - PubMed
-
- Goldstein JL, DeBose-Boyd RA, Brown MS (2006) Protein sensors for membrane sterols. Cell 124: 35–46. - PubMed
-
- Shen J, Chen X, Hendershot L, Prywes R (2002) ER stress regulation of ATF6 localization by dissociation of BiP/GRP78 binding and unmasking of Golgi localization signals. Dev Cell 3: 99–111. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
