

MYRF is a membrane-associated transcription factor that autoproteolytically cleaves to directly activate myelin genes
- PMID: 23966833
- PMCID: PMC3742440
- DOI: 10.1371/journal.pbio.1001625
MYRF is a membrane-associated transcription factor that autoproteolytically cleaves to directly activate myelin genes
Abstract
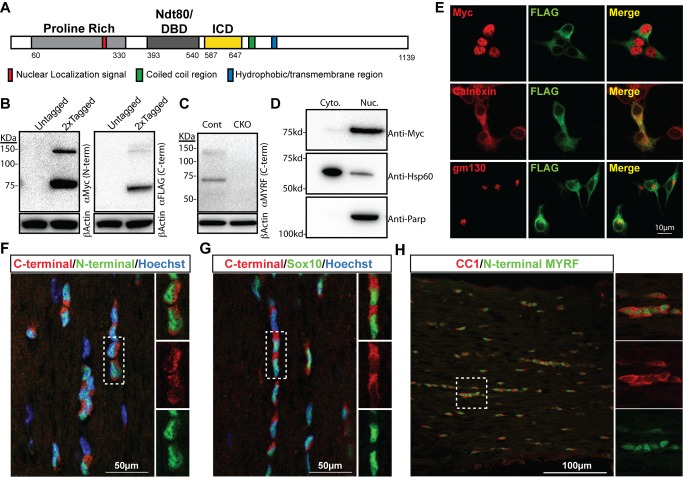
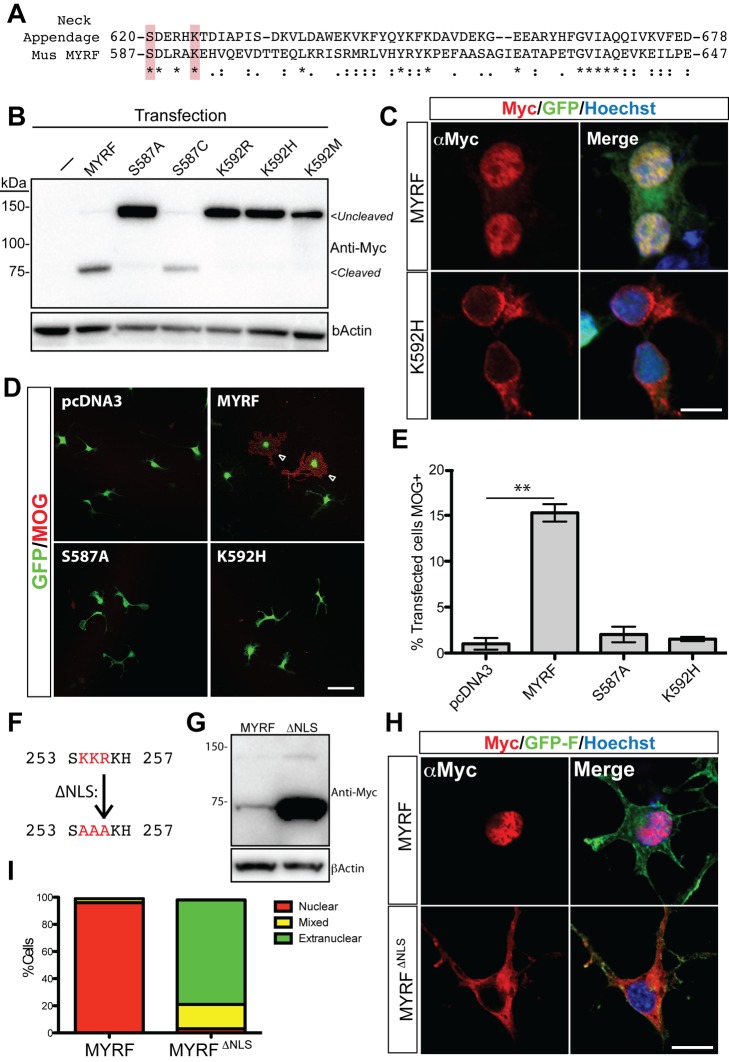
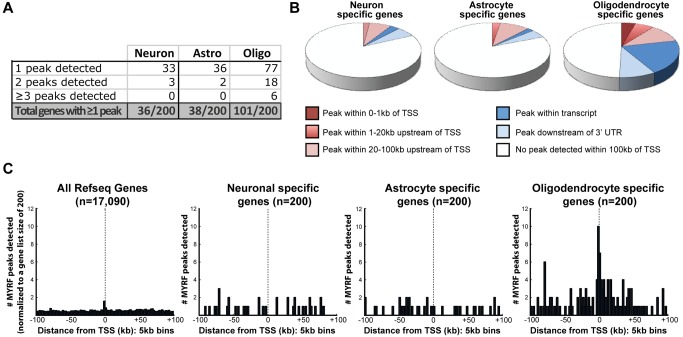
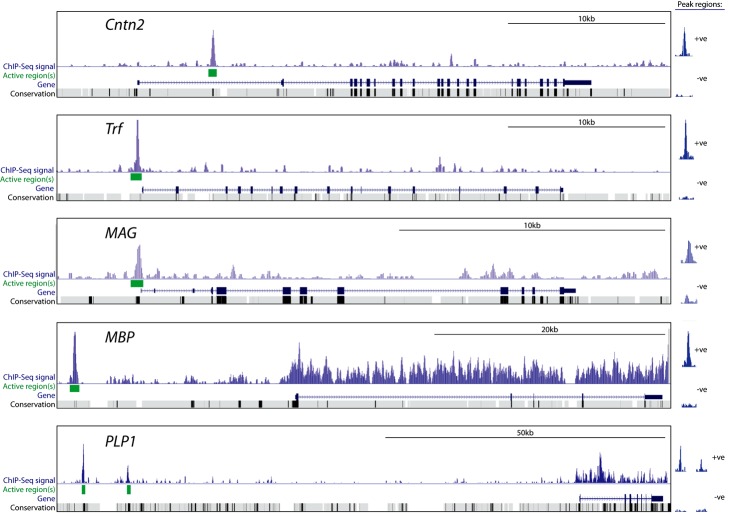
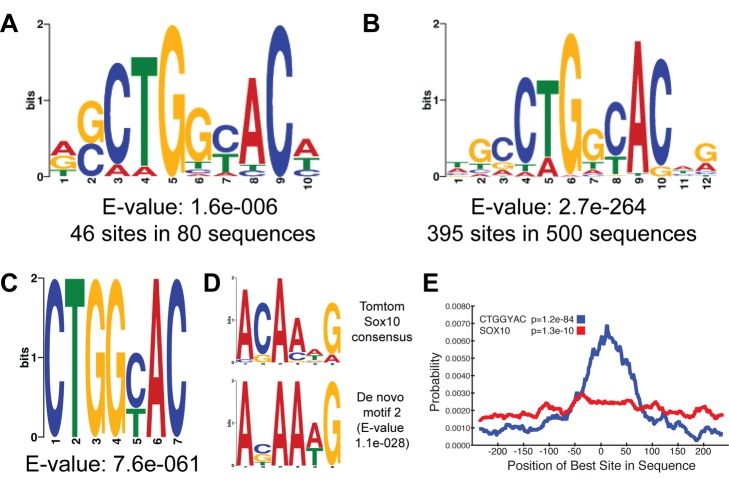
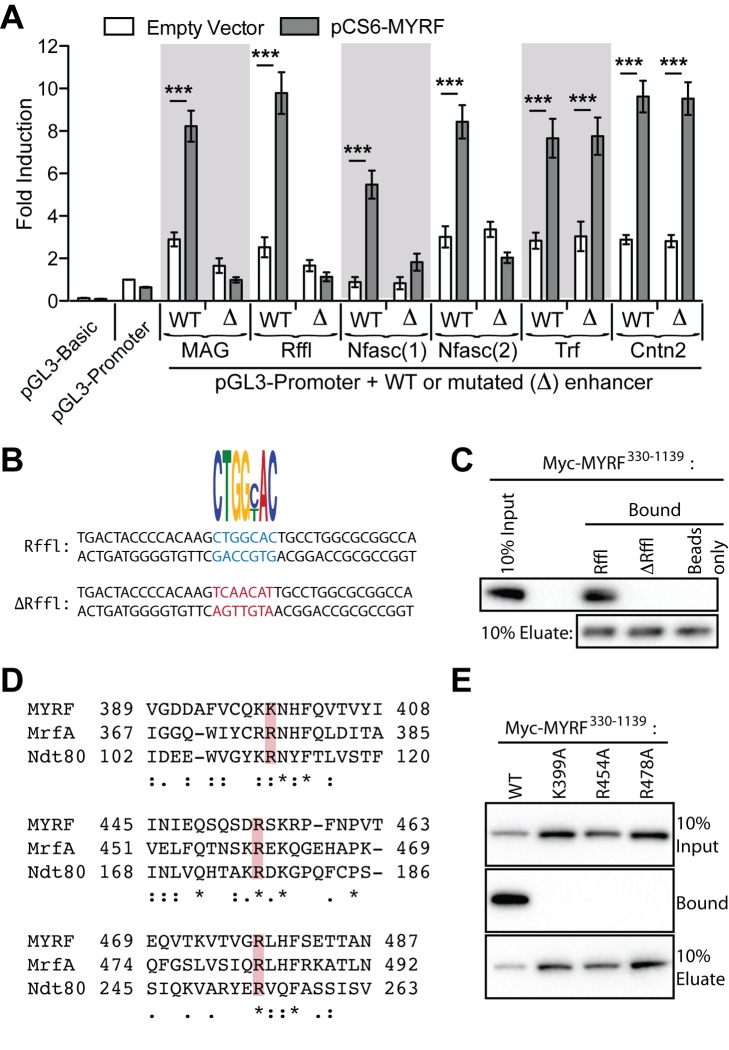
The myelination of axons is a crucial step during vertebrate central nervous system (CNS) development, allowing for rapid and energy efficient saltatory conduction of nerve impulses. Accordingly, the differentiation of oligodendrocytes, the myelinating cells of the CNS, and their expression of myelin genes are under tight transcriptional control. We previously identified a putative transcription factor, Myelin Regulatory Factor (Myrf), as being vital for CNS myelination. Myrf is required for the generation of CNS myelination during development and also for its maintenance in the adult. It has been controversial, however, whether Myrf directly regulates transcription, with reports of a transmembrane domain and lack of nuclear localization. Here we show that Myrf is a membrane-associated transcription factor that undergoes an activating proteolytic cleavage to separate its transmembrane domain-containing C-terminal region from a nuclear-targeted N-terminal region. Unexpectedly, this cleavage event occurs via a protein domain related to the autoproteolytic intramolecular chaperone domain of the bacteriophage tail spike proteins, the first time this domain has been found to play a role in eukaryotic proteins. Using ChIP-Seq we show that the N-terminal cleavage product directly binds the enhancer regions of oligodendrocyte-specific and myelin genes. This binding occurs via a defined DNA-binding consensus sequence and strongly promotes the expression of target genes. These findings identify Myrf as a novel example of a membrane-associated transcription factor and provide a direct molecular mechanism for its regulation of oligodendrocyte differentiation and CNS myelination.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

Comment in
-
Myelination borrows a trick from phage.PLoS Biol. 2013;11(8):e1001626. doi: 10.1371/journal.pbio.1001626. Epub 2013 Aug 13. PLoS Biol. 2013. PMID: 23966834 Free PMC article. No abstract available.
References
-
- Lu QR, Sun T, Zhu Z, Ma N, Garcia M, et al. (2002) Common developmental requirement for Olig function indicates a motor neuron/oligodendrocyte connection. Cell 109: 75–86. - PubMed
-
- Zhou Q, Anderson DJ (2002) The bHLH transcription factors OLIG2 and OLIG1 couple neuronal and glial subtype specification. Cell 109: 61–73. - PubMed
-
- Qi Y, Cai J, Wu Y, Wu R, Lee J, et al. (2001) Control of oligodendrocyte differentiation by the Nkx2.2 homeodomain transcription factor. Development 128: 2723–2733. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
