

The smallest capsid protein mediates binding of the essential tegument protein pp150 to stabilize DNA-containing capsids in human cytomegalovirus
- PMID: 23966856
- PMCID: PMC3744435
- DOI: 10.1371/journal.ppat.1003525
The smallest capsid protein mediates binding of the essential tegument protein pp150 to stabilize DNA-containing capsids in human cytomegalovirus
Abstract
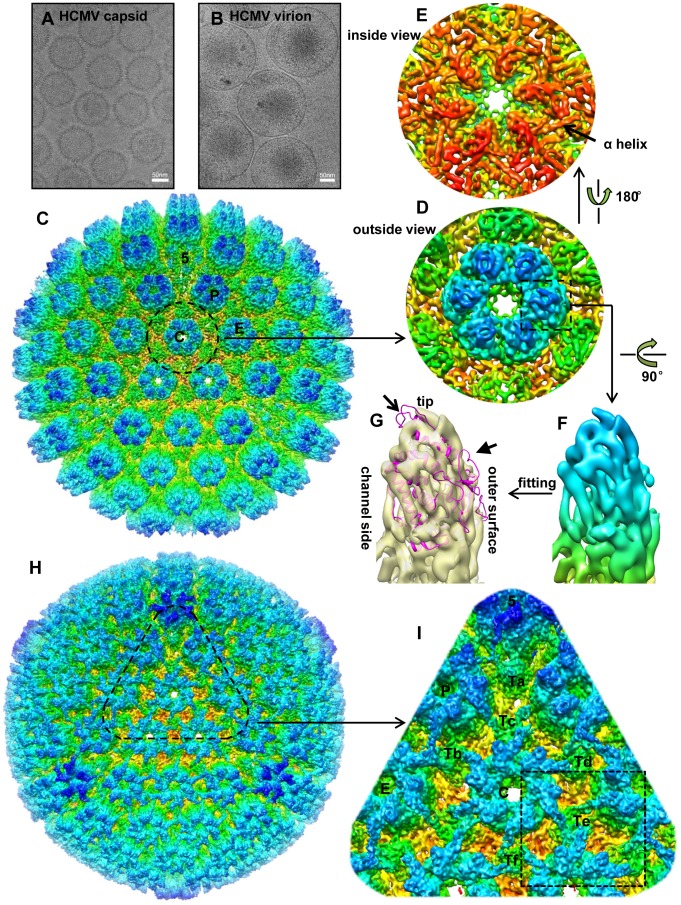
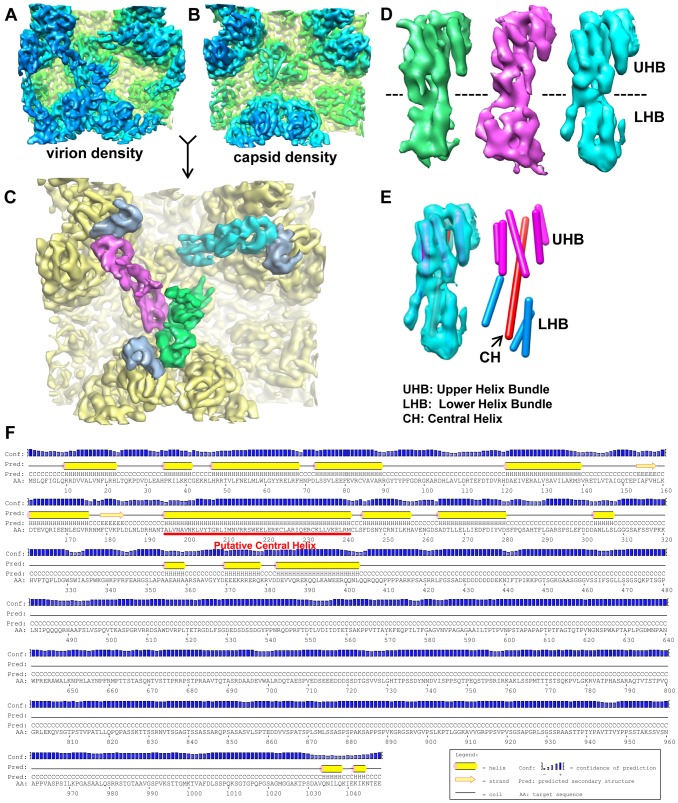
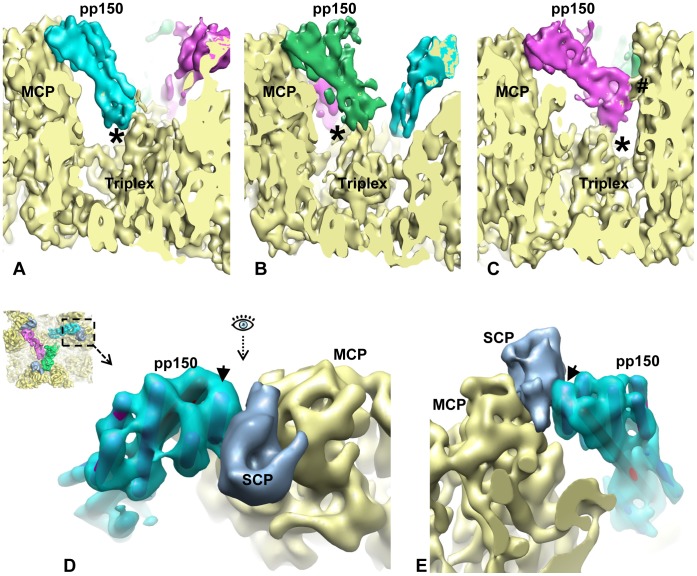
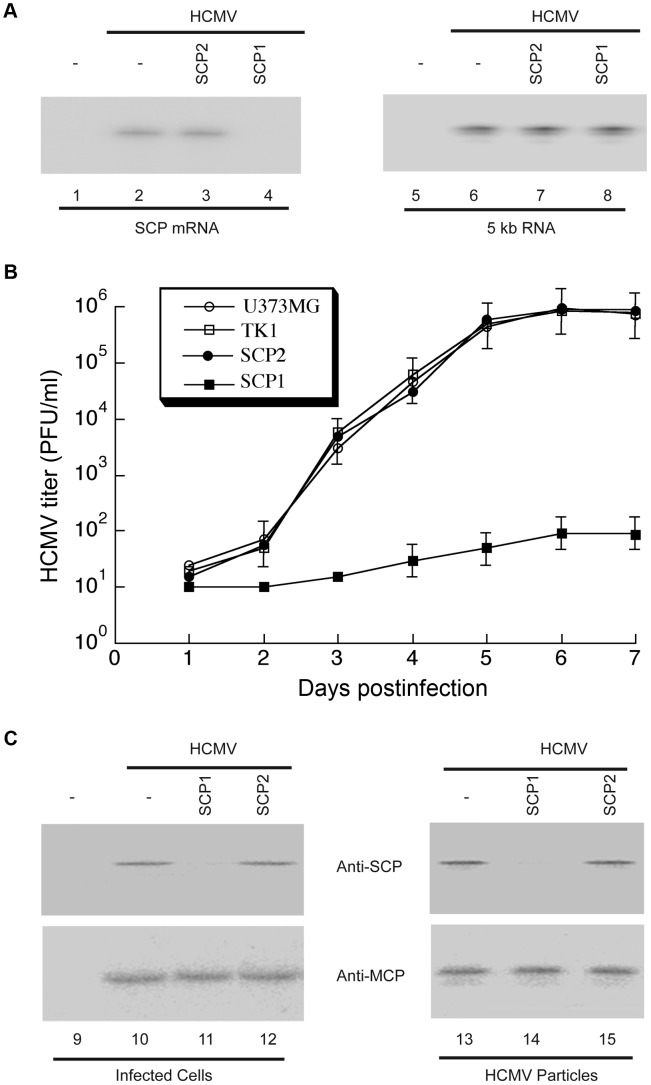
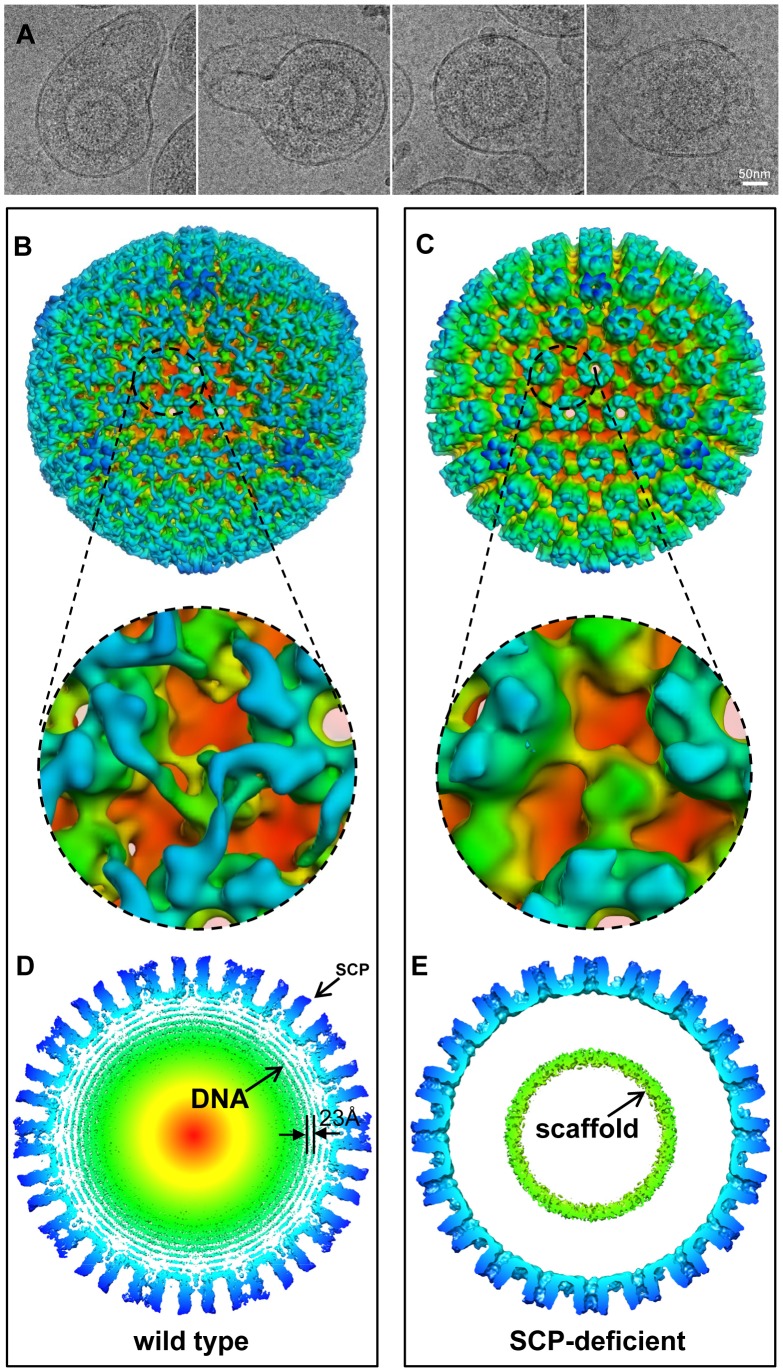
Human cytomegalovirus (HCMV) is a ubiquitous herpesvirus that causes birth defects in newborns and life-threatening complications in immunocompromised individuals. Among all human herpesviruses, HCMV contains a much larger dsDNA genome within a similarly-sized capsid compared to the others, and it was proposed to require pp150, a tegument protein only found in cytomegaloviruses, to stabilize its genome-containing capsid. However, little is known about how pp150 interacts with the underlying capsid. Moreover, the smallest capsid protein (SCP), while dispensable in herpes simplex virus type 1, was shown to play essential, yet undefined, role in HCMV infection. Here, by cryo electron microscopy (cryoEM), we determine three-dimensional structures of HCMV capsid (no pp150) and virion (with pp150) at sub-nanometer resolution. Comparison of these two structures reveals that each pp150 tegument density is composed of two helix bundles connected by a long central helix. Correlation between the resolved helices and sequence-based secondary structure prediction maps the tegument density to the N-terminal half of pp150. The structures also show that SCP mediates interactions between the capsid and pp150 at the upper helix bundle of pp150. Consistent with this structural observation, ribozyme inhibition of SCP expression in HCMV-infected cells impairs the formation of DNA-containing viral particles and reduces viral yield by 10,000 fold. By cryoEM reconstruction of the resulting "SCP-deficient" viral particles, we further demonstrate that SCP is required for pp150 functionally binding to the capsid. Together, our structural and biochemical results point to a mechanism whereby SCP recruits pp150 to stabilize genome-containing capsid for the production of infectious HCMV virion.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Mocarski ES, Shenk T, Pass RF (2007) Cytomegalovirus. In: Knipe DM, Howley PM, Griffin DE, Martin MA, Lamb RA et al.., editors. Fields Virology. Philadelphia, PA: Lippincott/Williams & Wilkins. pp. 2701–2771.
-
- Britt WJ, Boppana S (2004) Human cytomegalovirus virion proteins. Hum Immunol 65: 395–402. - PubMed
