

Eat to reproduce: a key role for the insulin signaling pathway in adult insects
- PMID: 23966944
- PMCID: PMC3735985
- DOI: 10.3389/fphys.2013.00202
Eat to reproduce: a key role for the insulin signaling pathway in adult insects
Abstract
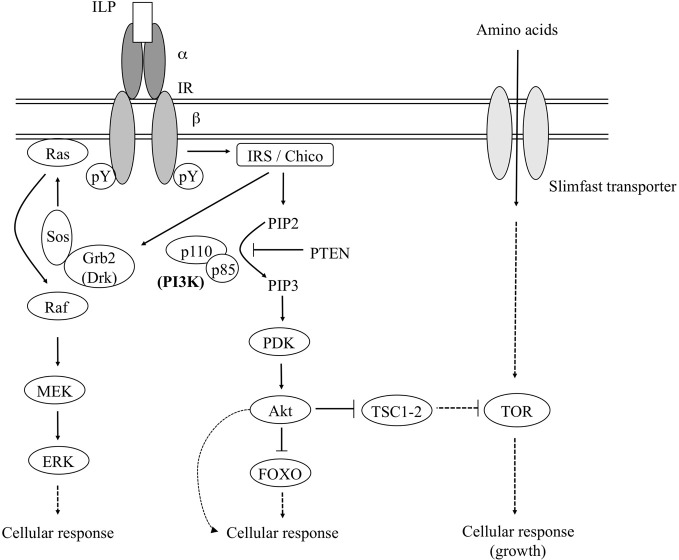
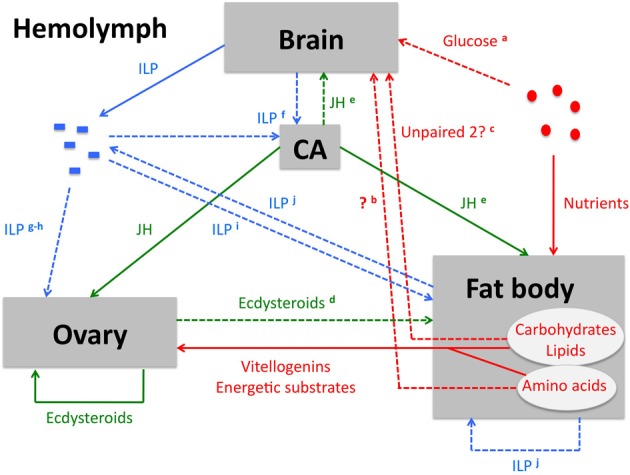
Insects, like all heterotrophic organisms, acquire from their food the nutrients that are essential for anabolic processes that lead to growth (larval stages) or reproduction (adult stage). In adult females, this nutritional input is processed and results in a very specific output, i.e., the production of fully developed eggs ready for fertilization and deposition. An important role in this input-output transition is attributed to the insulin signaling pathway (ISP). The ISP is considered to act as a sensor of the organism's nutritional status and to stimulate the progression of anabolic events when the status is positive. In several insect species belonging to different orders, the ISP has been demonstrated to positively control vitellogenesis and oocyte growth. Whether or not ISP acts herein via a mediator action of lipophilic insect hormones (ecdysteroids and juvenile hormone) remains debatable and might be differently controlled in different insect orders. Most likely, insulin-related peptides, ecdysteroids and juvenile hormone are involved in a complex regulatory network, in which they mutually influence each other and in which the insect's nutritional status is a crucial determinant of the network's output. The current review will present an overview of the regulatory role of the ISP in female insect reproduction and its interaction with other pathways involving nutrients, lipophilic hormones and neuropeptides.
Keywords: female insect reproduction; insulin signaling pathway; lipophilic hormones; neuropeptides; nutritional status.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources