

Albumin is recycled from the primary urine by tubular transcytosis
- PMID: 23970123
- PMCID: PMC3839546
- DOI: 10.1681/ASN.2013010018
Albumin is recycled from the primary urine by tubular transcytosis
Abstract
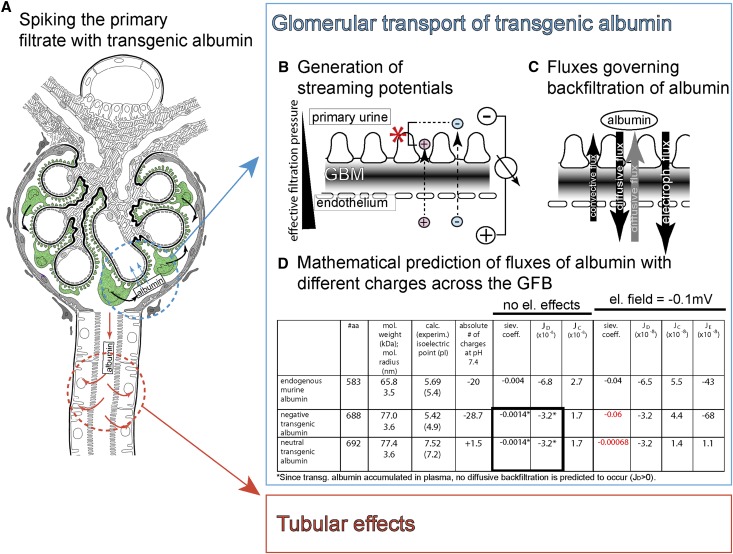
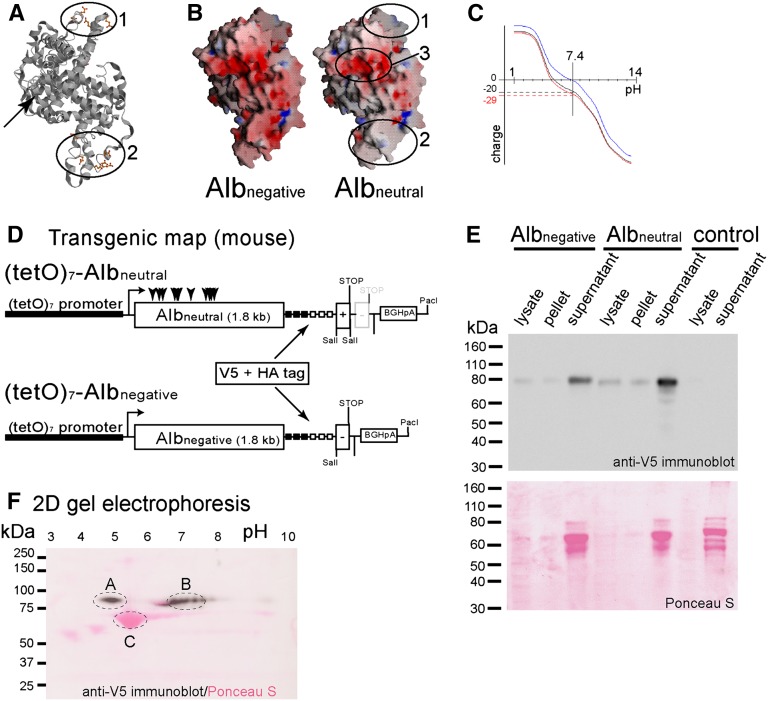
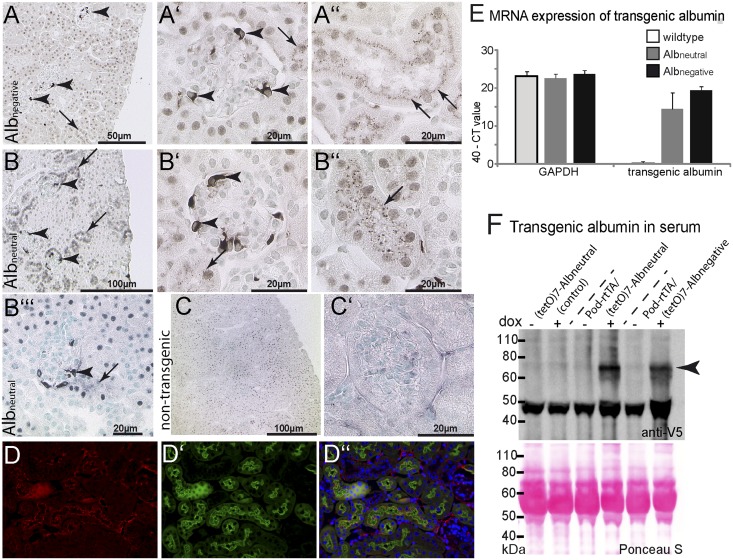
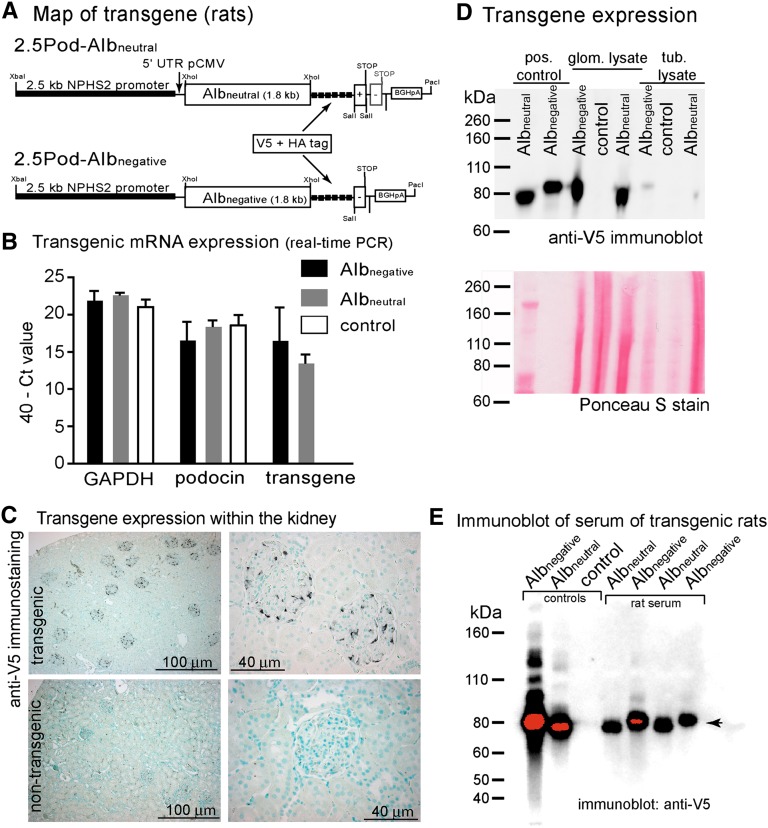
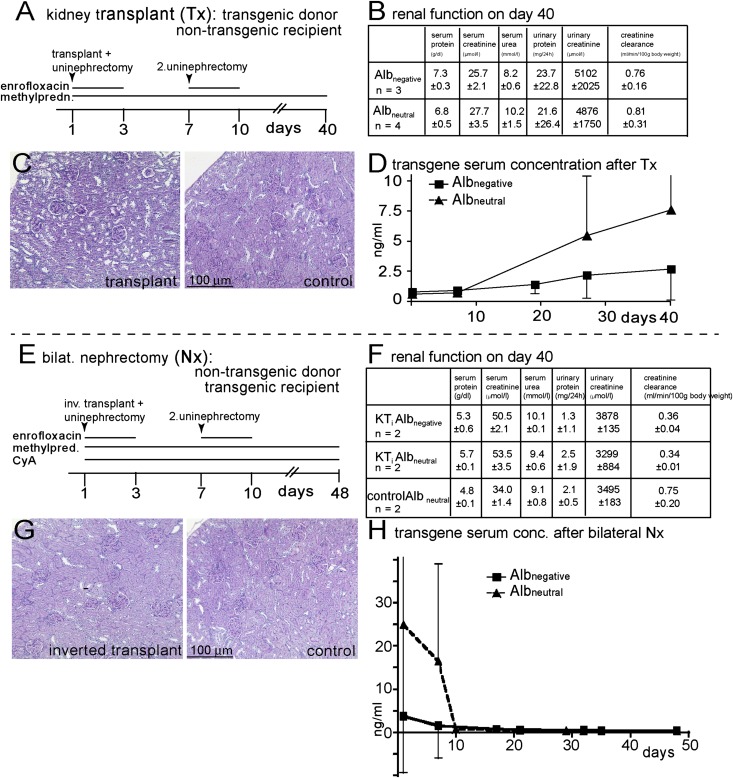
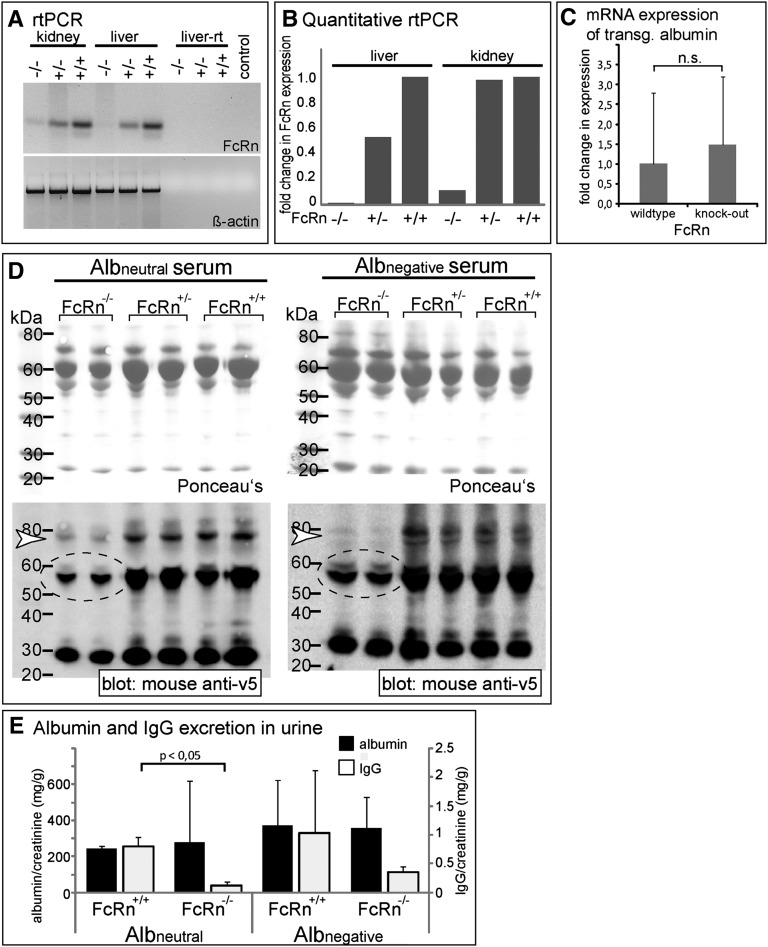
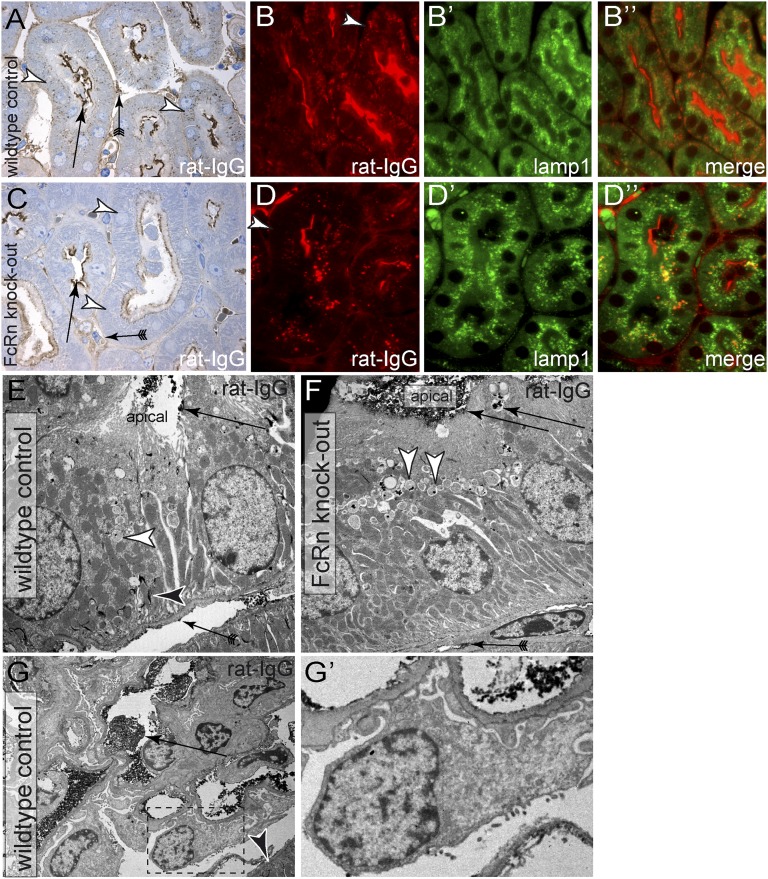
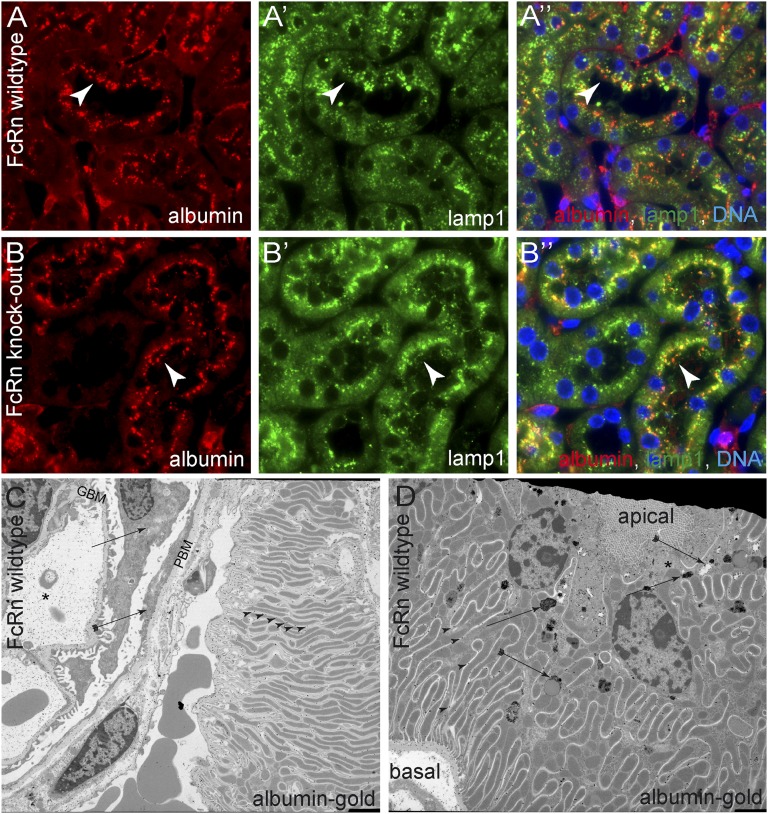
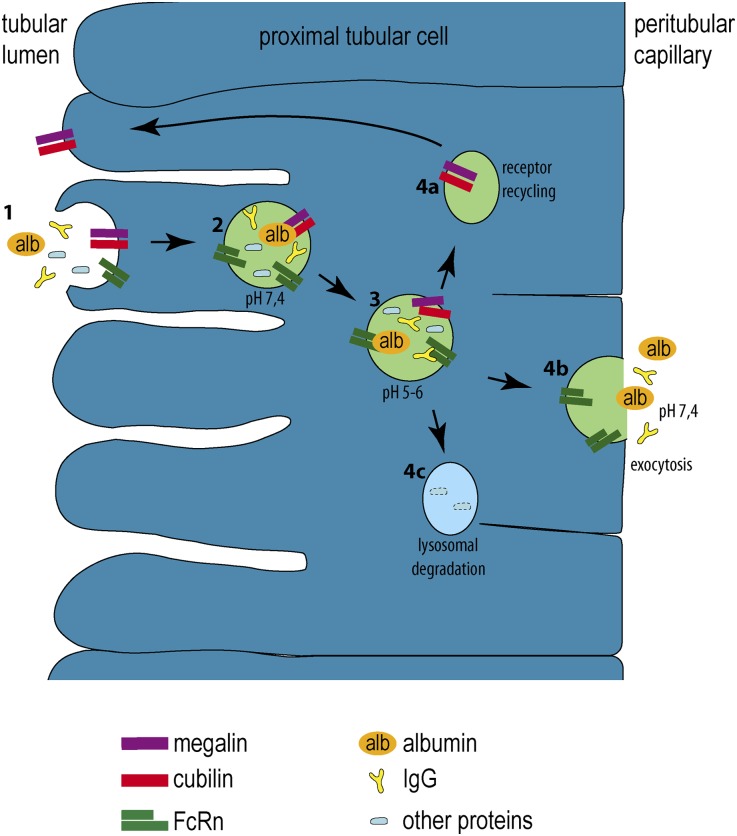
Under physiologic conditions, significant amounts of plasma protein pass the renal filter and are reabsorbed by proximal tubular cells, but it is not clear whether the endocytosed protein, particularly albumin, is degraded in lysosomes or returned to the circulatory system intact. To resolve this question, a transgenic mouse with podocyte-specific expression of doxycycline-inducible tagged murine albumin was developed. To assess potential glomerular backfiltration, two types of albumin with different charges were expressed. On administration of doxycycline, podocytes expressed either of the two types of transgenic albumin, which were secreted into the primary filtrate and reabsorbed by proximal tubular cells, resulting in serum accumulation. Renal transplantation experiments confirmed that extrarenal transcription of transgenic albumin was unlikely to account for these results. Genetic deletion of the neonatal Fc receptor (FcRn), which rescues albumin and IgG from lysosomal degradation, abolished transcytosis of both types of transgenic albumin and IgG in proximal tubular cells. In summary, we provide evidence of a transcytosis within the kidney tubular system that protects albumin and IgG from lysosomal degradation, allowing these proteins to be recycled intact.
Figures
Comment in
-
Proteinuria: Tubular handling of albumin-degradation or salvation?Nat Rev Nephrol. 2013 Dec;9(12):700-2. doi: 10.1038/nrneph.2013.212. Epub 2013 Oct 15. Nat Rev Nephrol. 2013. PMID: 24126590
References
-
- Peters T, Jr: Serum albumin: Recent progress in the understanding of its structure and biosynthesis. Clin Chem 23: 5–12, 1977. - PubMed
-
- Haraldsson B, Nyström J, Deen WM: Properties of the glomerular barrier and mechanisms of proteinuria. Physiol Rev 88: 451–487, 2008. - PubMed
-
- Tojo A, Endou H: Intrarenal handling of proteins in rats using fractional micropuncture technique. Am J Physiol 263: F601–F606, 1992. - PubMed
-
- Norden AG, Lapsley M, Lee PJ, Pusey CD, Scheinman SJ, Tam FW, Thakker RV, Unwin RJ, Wrong O: Glomerular protein sieving and implications for renal failure in Fanconi syndrome. Kidney Int 60: 1885–1892, 2001. - PubMed
-
- Christensen EI, Birn H, Rippe B, Maunsbach AB: Controversies in nephrology: Renal albumin handling, facts, and artifacts! Kidney Int 72: 1192–1194, 2007. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
