

Enhanced cytotoxicity from deoxyguanosine-enriched T-oligo in prostate cancer cells
- PMID: 23971906
- PMCID: PMC3760086
- DOI: 10.1089/nat.2013.0420
Enhanced cytotoxicity from deoxyguanosine-enriched T-oligo in prostate cancer cells
Abstract
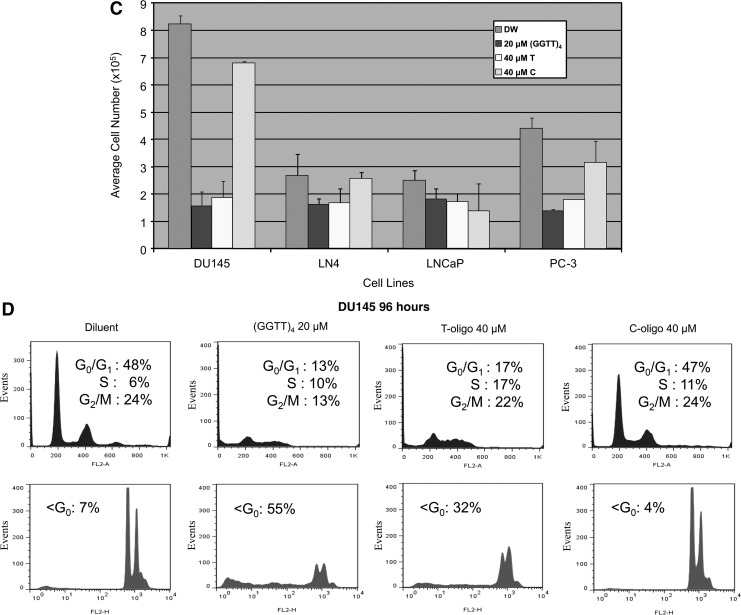
Prostate cancer represents approximately 10 percent of all cancer cases in men and accounts for more than a quarter of all cancer types. Advances in understanding the molecular mechanisms of prostate cancer progression, however, have not translated well to the clinic. Patients with metastatic and hormone-refractory disease have only palliative options for treatment, as chemotherapy seldom produces durable or complete responses, highlighting the need for novel therapeutic approaches. T-oligo, a single-stranded deoxyribonucleic acid with partial sequence homology to human telomeric DNA, has elicited cytostatic and/or cytotoxic effects in multiple cancer cell types. In contrast, normal primary cells of varying tissue types are resistant to cytotoxic actions of T-oligo, underscoring its potential utility as a novel targeted cancer therapeutic. Mechanistically, T-oligo is hypothesized to interfere with normal telomeric structure and form G-quadruplex structures, thereby inducing genomic stress in addition to aberrant upregulation of DNA damageresponse pathways. Here, we present data demonstrating the enhanced effectiveness of a deoxyguanosine-enriched sequence of T-oligo, termed (GGTT)4, which elicits robust cytotoxic effects in prostate cancer cells at lower concentrations than the most recent T-oligo sequence (5'-pGGT TAG GTG TAG GTT T 3') described to date and used for comparison in this study, while exerting no cytotoxic actions on nontransformed human prostate epithelial cells. Additionally, we provide evidence supporting the T-oligo induced activation of cJun N-terminal kinase (JNK) signaling in prostate cancer cells consistent with G-quadruplex formation, thereby significantly advancing the understanding of the T-oligo mechanism of action.
Figures







References
-
- AOKI H. IWADO E. ELLER M.S. KONDO Y. FUJIWARA K. LI G.Z. HESS K.R. SIWAK D.R. SAWAYA R. MILLS G.B., et al. Telomere 3′ overhang-specific DNA oligonucleotides induce autophagy in malignant glioma cells. FASEB J. 2007;21:2918–2930. - PubMed
-
- ELLER M.S. PURI N. HADSHIEW I.M. VENNA S.S. GILCHREST B.A. Induction of apoptosis by telomere 3′ overhang specific DNA. Exp. Cell Res. 2002;276:185–193. - PubMed
-
- ELLER M.S. LI G.Z. FIROOZABADI R. PURI N. GILCHREST B.A. Induction of a p95/Nbs-1- mediated S phase checkpoint by telomere 3′ overhang specific DNA. FASEB J. 2003;2:152–162. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
