

Learning from the spinal cord: how the study of spinal cord plasticity informs our view of learning
- PMID: 23973905
- PMCID: PMC3946174
- DOI: 10.1016/j.nlm.2013.08.003
Learning from the spinal cord: how the study of spinal cord plasticity informs our view of learning
Abstract
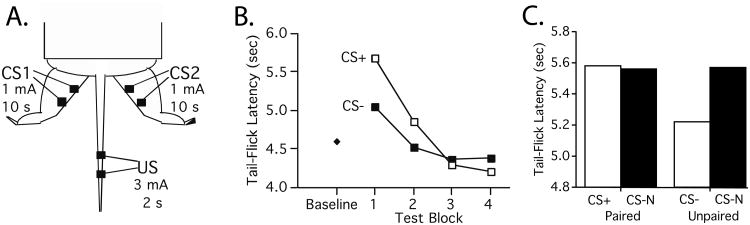
The paper reviews research examining whether and how training can induce a lasting change in spinal cord function. A framework for the study of learning, and some essential issues in experimental design, are discussed. A core element involves delayed assessment under common conditions. Research has shown that brain systems can induce a lasting (memory-like) alteration in spinal function. Neurons within the lower (lumbosacral) spinal cord can also adapt when isolated from the brain by means of a thoracic transection. Using traditional learning paradigms, evidence suggests that spinal neurons support habituation and sensitization as well as Pavlovian and instrumental conditioning. At a neurobiological level, spinal systems support phenomena (e.g., long-term potentiation), and involve mechanisms (e.g., NMDA mediated plasticity, protein synthesis) implicated in brain-dependent learning and memory. Spinal learning also induces modulatory effects that alter the capacity for learning. Uncontrollable/unpredictable stimulation disables the capacity for instrumental learning and this effect has been linked to the cytokine tumor necrosis factor (TNF). Predictable/controllable stimulation enables learning and counters the adverse effects of uncontrollable stimulation through a process that depends upon brain-derived neurotrophic factor (BDNF). Finally, uncontrollable, but not controllable, nociceptive stimulation impairs recovery after a contusion injury. A process-oriented approach (neurofunctionalism) is outlined that encourages a broader view of learning phenomena.
Keywords: Inflammation; Injury; Instrumental conditioning; Operant; Pavlovian conditioning; Spinal cord.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures






Similar articles
-
Metaplasticity and behavior: how training and inflammation affect plastic potential within the spinal cord and recovery after injury.Front Neural Circuits. 2014 Sep 8;8:100. doi: 10.3389/fncir.2014.00100. eCollection 2014. Front Neural Circuits. 2014. PMID: 25249941 Free PMC article. Review.
-
Metaplasticity within the spinal cord: Evidence brain-derived neurotrophic factor (BDNF), tumor necrosis factor (TNF), and alterations in GABA function (ionic plasticity) modulate pain and the capacity to learn.Neurobiol Learn Mem. 2018 Oct;154:121-135. doi: 10.1016/j.nlm.2018.04.007. Epub 2018 Apr 7. Neurobiol Learn Mem. 2018. PMID: 29635030 Free PMC article.
-
Instrumental learning within the spinal cord: underlying mechanisms and implications for recovery after injury.Behav Cogn Neurosci Rev. 2006 Dec;5(4):191-239. doi: 10.1177/1534582306289738. Behav Cogn Neurosci Rev. 2006. PMID: 17099112 Review.
-
Brain-derived neurotrophic factor promotes adaptive plasticity within the spinal cord and mediates the beneficial effects of controllable stimulation.Neuroscience. 2012 Jan 3;200:74-90. doi: 10.1016/j.neuroscience.2011.10.028. Epub 2011 Oct 25. Neuroscience. 2012. PMID: 22056599 Free PMC article.
-
Learning to promote recovery after spinal cord injury.Exp Neurol. 2020 Aug;330:113334. doi: 10.1016/j.expneurol.2020.113334. Epub 2020 Apr 28. Exp Neurol. 2020. PMID: 32353465 Free PMC article. Review.
Cited by
-
Operant conditioning of spinal reflexes: from basic science to clinical therapy.Front Integr Neurosci. 2014 Mar 18;8:25. doi: 10.3389/fnint.2014.00025. eCollection 2014. Front Integr Neurosci. 2014. PMID: 24672441 Free PMC article. Review.
-
Metaplasticity and behavior: how training and inflammation affect plastic potential within the spinal cord and recovery after injury.Front Neural Circuits. 2014 Sep 8;8:100. doi: 10.3389/fncir.2014.00100. eCollection 2014. Front Neural Circuits. 2014. PMID: 25249941 Free PMC article. Review.
-
The left-right side-specific endocrine signaling in the effects of brain lesions: questioning of the neurological dogma.Cell Mol Life Sci. 2022 Oct 11;79(11):545. doi: 10.1007/s00018-022-04576-9. Cell Mol Life Sci. 2022. PMID: 36219330 Free PMC article. Review.
-
Simultaneous Brain-Cervical Cord fMRI Reveals Intrinsic Spinal Cord Plasticity during Motor Sequence Learning.PLoS Biol. 2015 Jun 30;13(6):e1002186. doi: 10.1371/journal.pbio.1002186. eCollection 2015 Jun. PLoS Biol. 2015. PMID: 26125597 Free PMC article.
-
Evidence That the Central Nervous System Can Induce a Modification at the Neuromuscular Junction That Contributes to the Maintenance of a Behavioral Response.J Neurosci. 2020 Nov 25;40(48):9186-9209. doi: 10.1523/JNEUROSCI.2683-19.2020. Epub 2020 Oct 23. J Neurosci. 2020. PMID: 33097637 Free PMC article.
References
-
- Abraham WC, Bear MF. Metaplasticity: the plasticity of synaptic plasticity. Trends in Neuroscience. 1996;19:126–130. - PubMed
-
- Allen C, Grau JW, Meagher MW. The lower bounds of cognition: What do spinal cords reveal? In: Bickle J, editor. The Oxford Handbook of Philosophy of Neuroscience. Oxford: Oxford Press; 2009. pp. 129–142.
-
- Bardin L, Schmidt J, Alloui A, Eschalier A. Effect of intrathecal administration of serotonin in chronic pain models in rats. European Journal of Pharmacology. 2000;409:37–43. - PubMed
-
- Basbaum AI, Fields HL. Endogenous pain control systems: Brainstem spinal pathways and endorphin circuitry. Annual Review of Neuroscience. 1984;7:309–338. - PubMed
-
- Basso DM, Beattie MS, Bresnahan JC. A sensitive and reliable locomotive rating scale for open field testing in rats. Journal of Neurotrauma. 1995;12:1–21. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
