

CAS directly interacts with vinculin to control mechanosensing and focal adhesion dynamics
- PMID: 23974298
- PMCID: PMC3901934
- DOI: 10.1007/s00018-013-1450-x
CAS directly interacts with vinculin to control mechanosensing and focal adhesion dynamics
Abstract
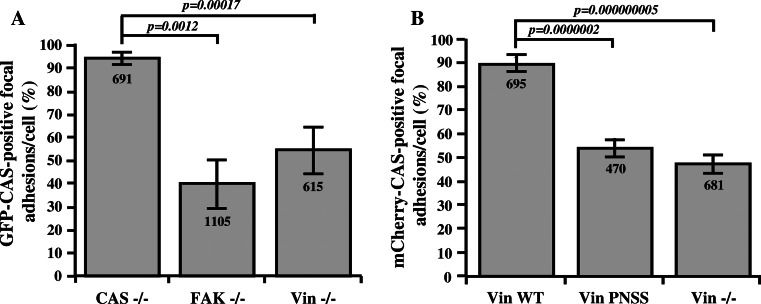
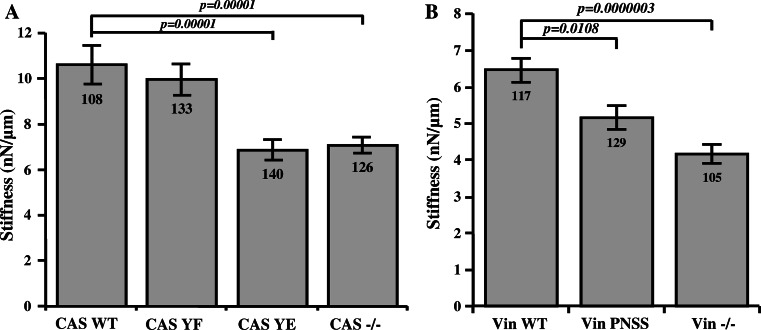
Focal adhesions are cellular structures through which both mechanical forces and regulatory signals are transmitted. Two focal adhesion-associated proteins, Crk-associated substrate (CAS) and vinculin, were both independently shown to be crucial for the ability of cells to transmit mechanical forces and to regulate cytoskeletal tension. Here, we identify a novel, direct binding interaction between CAS and vinculin. This interaction is mediated by the CAS SRC homology 3 domain and a proline-rich sequence in the hinge region of vinculin. We show that CAS localization in focal adhesions is partially dependent on vinculin, and that CAS-vinculin coupling is required for stretch-induced activation of CAS at the Y410 phosphorylation site. Moreover, CAS-vinculin binding significantly affects the dynamics of CAS and vinculin within focal adhesions as well as the size of focal adhesions. Finally, disruption of CAS binding to vinculin reduces cell stiffness and traction force generation. Taken together, these findings strongly implicate a crucial role of CAS-vinculin interaction in mechanosensing and focal adhesion dynamics.
Figures








References
-
- Dorssers LCJ, Grebenchtchikov N, Brinkman A, Look MP, Klijn AGM, Geurts-Moespot A, Span PN, Foekens JA, Sweep CGJ. Application of a newly developed ELISA for BCAR1 protein for prediction of clinical benefit of tamoxifen therapy in patients with advanced breast cancer. Clin Chem. 2004;50:1445–1447. doi: 10.1373/clinchem.2004.035493. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
