

Recent advances in 2D and 3D in vitro systems using primary hepatocytes, alternative hepatocyte sources and non-parenchymal liver cells and their use in investigating mechanisms of hepatotoxicity, cell signaling and ADME
- PMID: 23974980
- PMCID: PMC3753504
- DOI: 10.1007/s00204-013-1078-5
Recent advances in 2D and 3D in vitro systems using primary hepatocytes, alternative hepatocyte sources and non-parenchymal liver cells and their use in investigating mechanisms of hepatotoxicity, cell signaling and ADME
Abstract
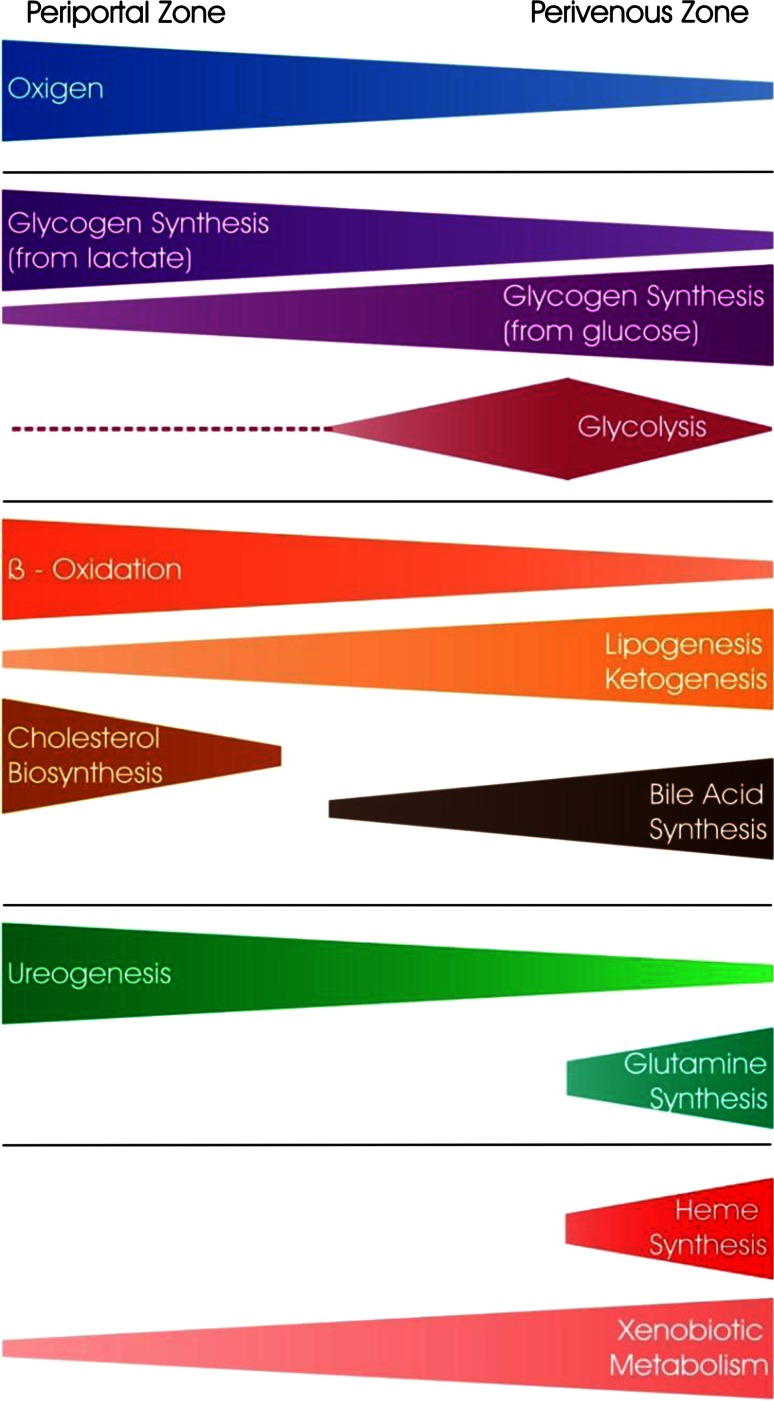
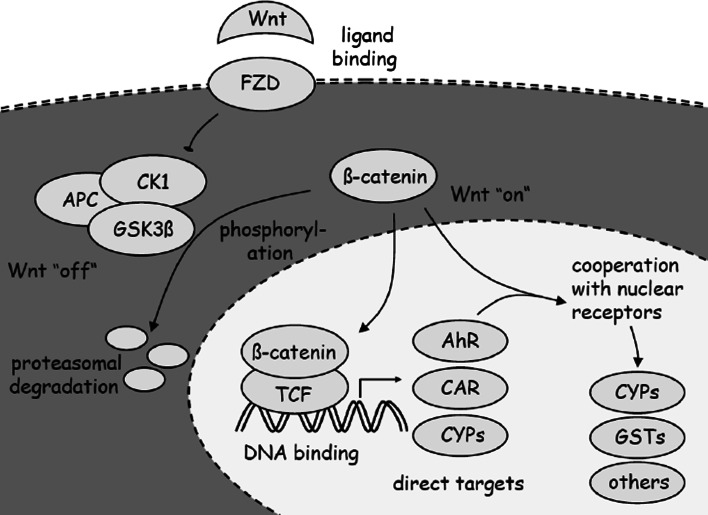
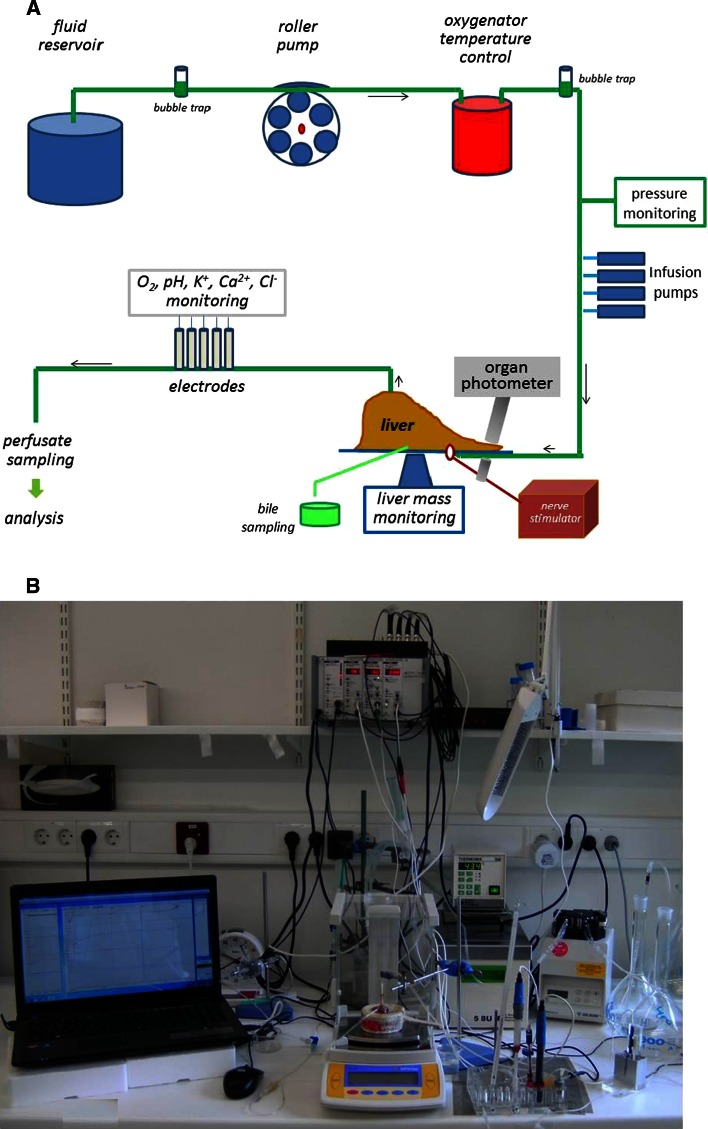
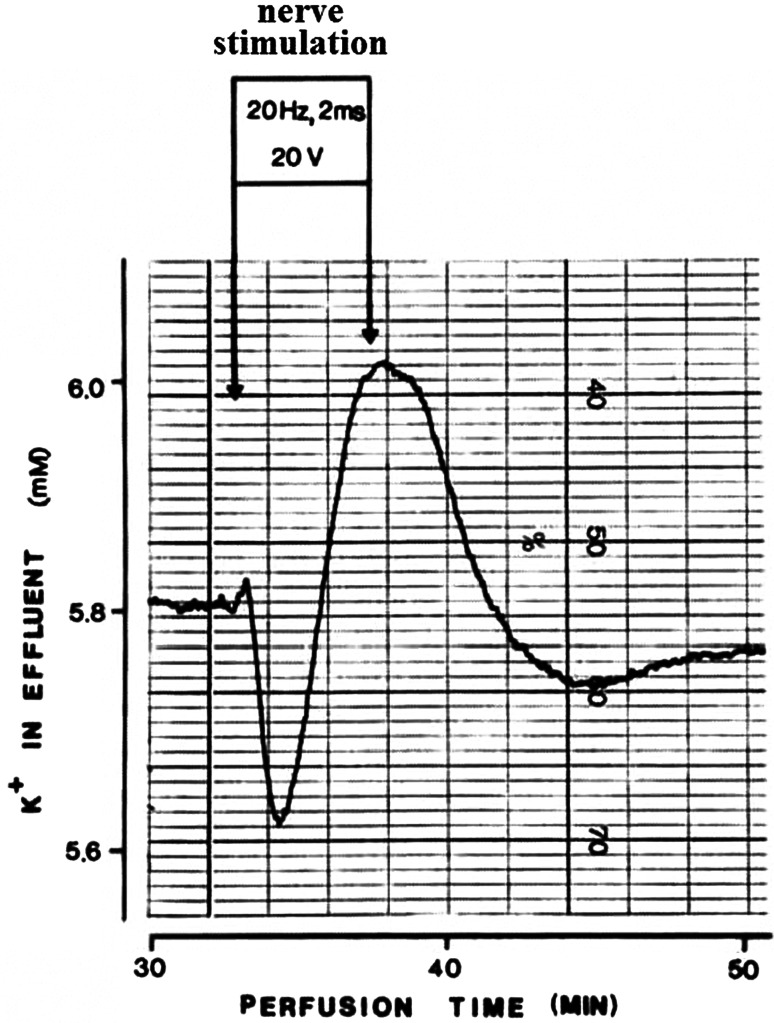
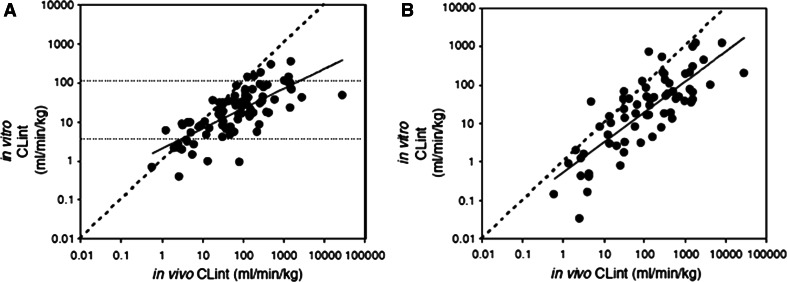
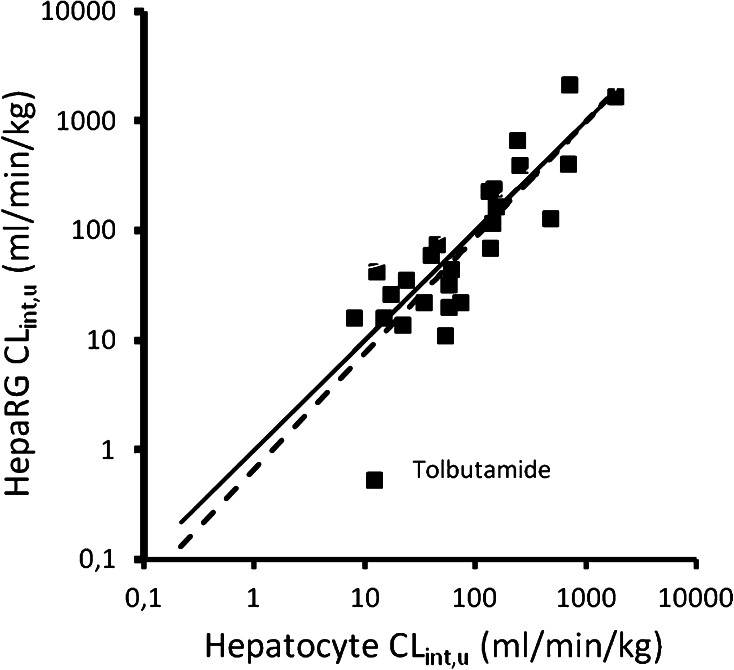
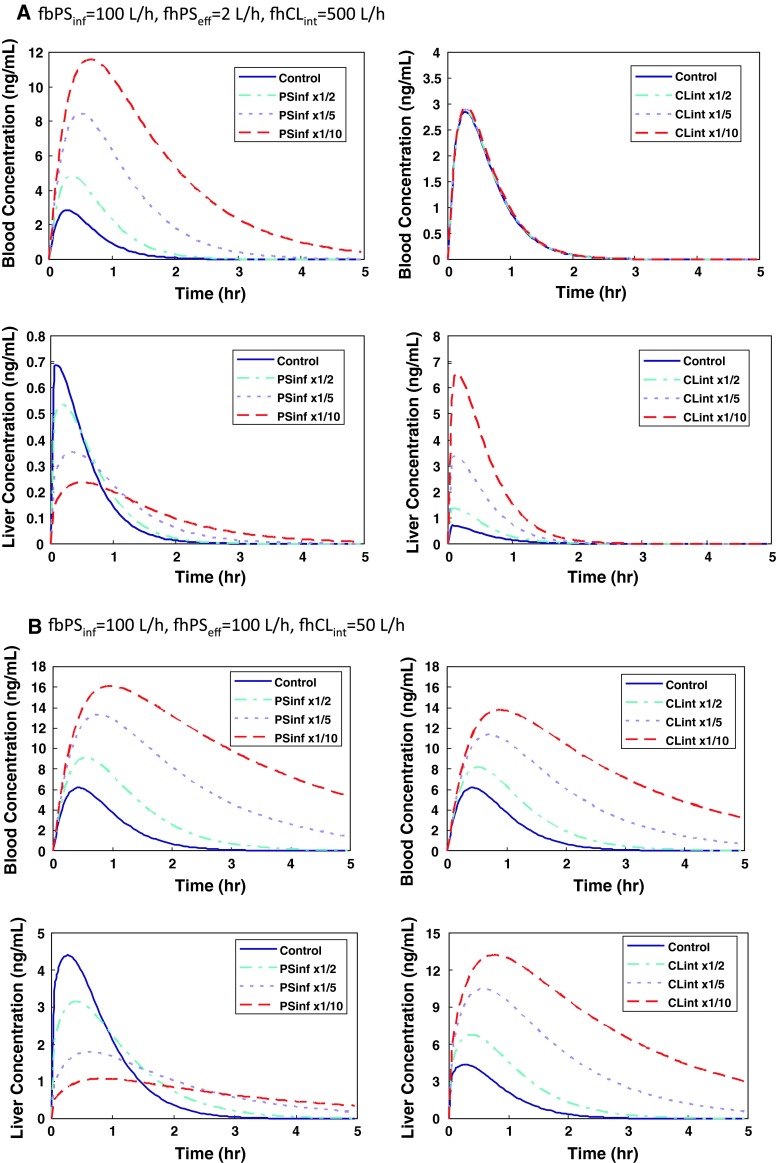
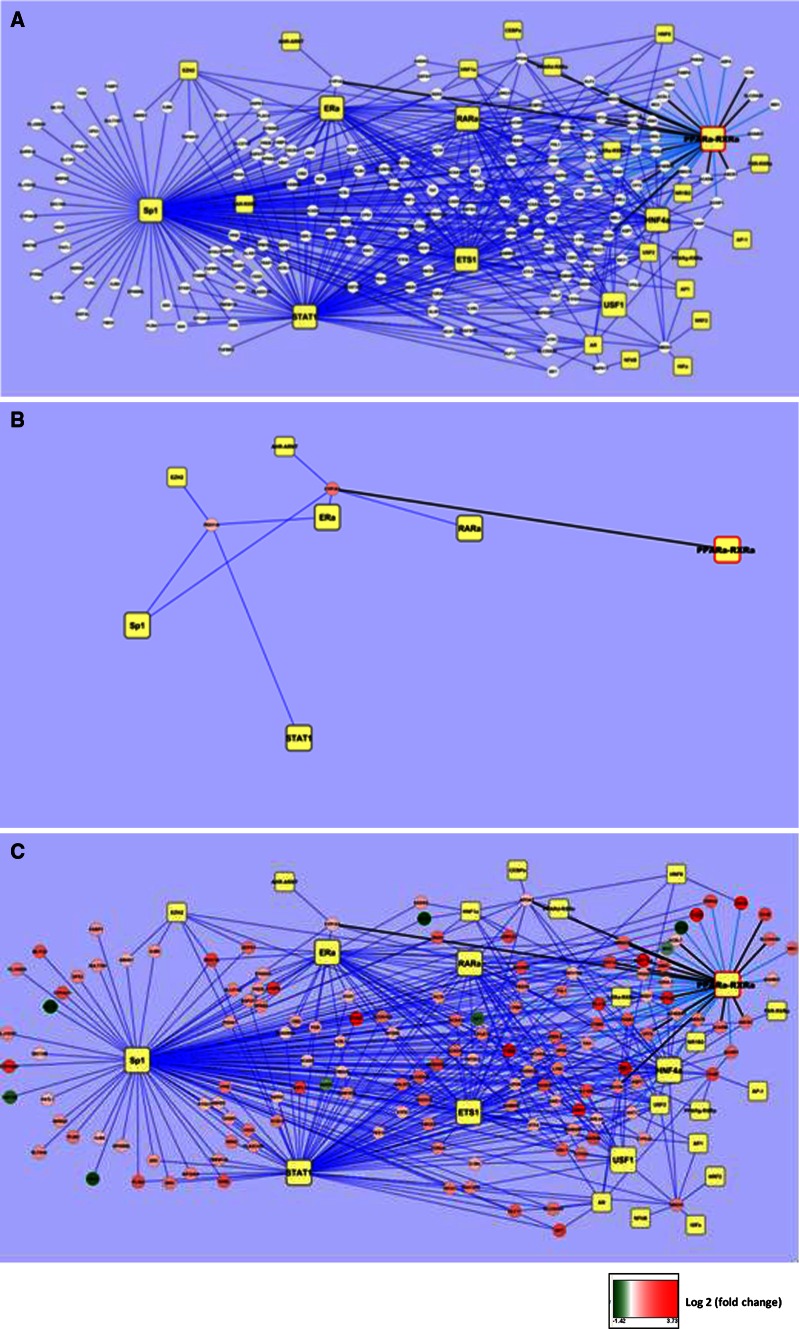
This review encompasses the most important advances in liver functions and hepatotoxicity and analyzes which mechanisms can be studied in vitro. In a complex architecture of nested, zonated lobules, the liver consists of approximately 80 % hepatocytes and 20 % non-parenchymal cells, the latter being involved in a secondary phase that may dramatically aggravate the initial damage. Hepatotoxicity, as well as hepatic metabolism, is controlled by a set of nuclear receptors (including PXR, CAR, HNF-4α, FXR, LXR, SHP, VDR and PPAR) and signaling pathways. When isolating liver cells, some pathways are activated, e.g., the RAS/MEK/ERK pathway, whereas others are silenced (e.g. HNF-4α), resulting in up- and downregulation of hundreds of genes. An understanding of these changes is crucial for a correct interpretation of in vitro data. The possibilities and limitations of the most useful liver in vitro systems are summarized, including three-dimensional culture techniques, co-cultures with non-parenchymal cells, hepatospheres, precision cut liver slices and the isolated perfused liver. Also discussed is how closely hepatoma, stem cell and iPS cell-derived hepatocyte-like-cells resemble real hepatocytes. Finally, a summary is given of the state of the art of liver in vitro and mathematical modeling systems that are currently used in the pharmaceutical industry with an emphasis on drug metabolism, prediction of clearance, drug interaction, transporter studies and hepatotoxicity. One key message is that despite our enthusiasm for in vitro systems, we must never lose sight of the in vivo situation. Although hepatocytes have been isolated for decades, the hunt for relevant alternative systems has only just begun.
Figures
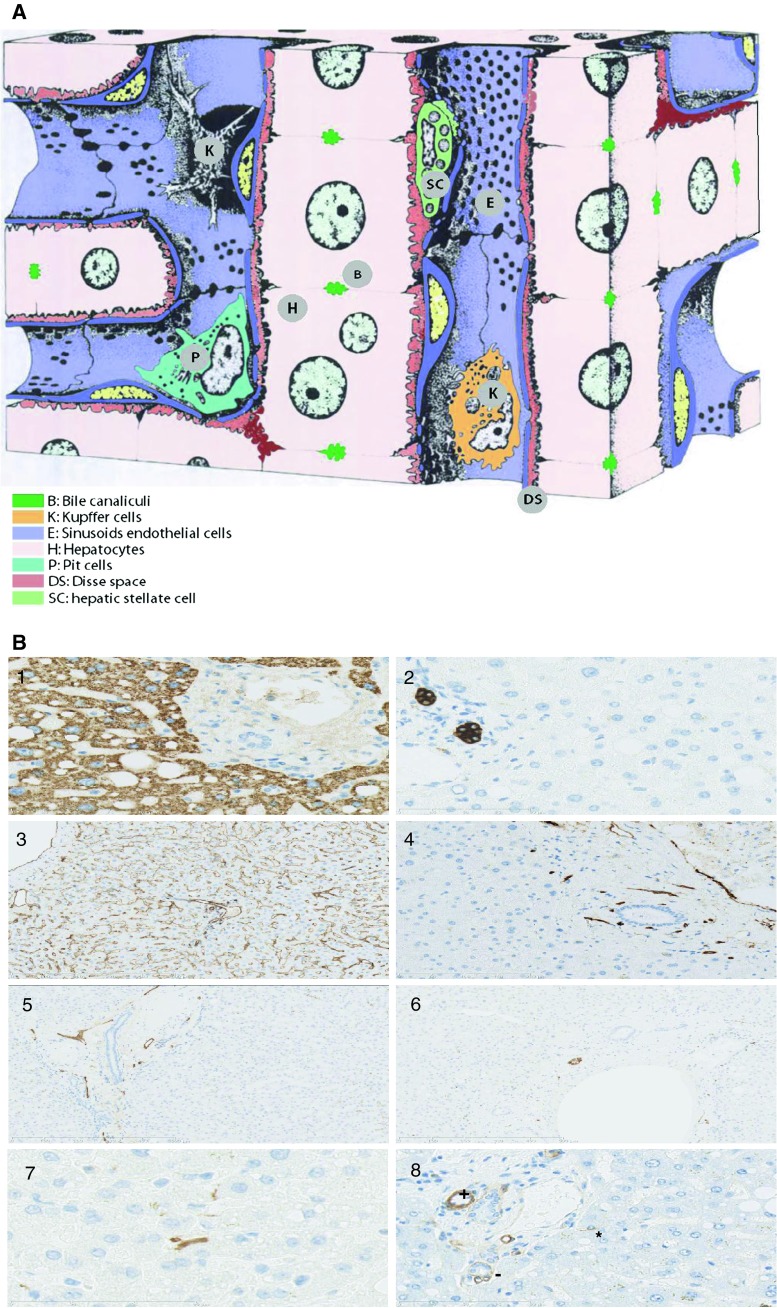
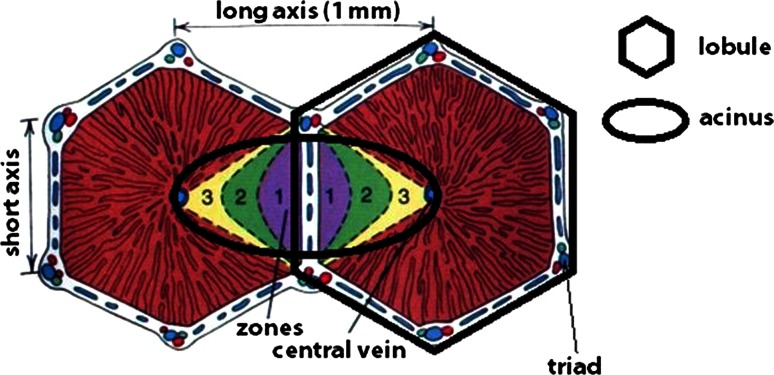
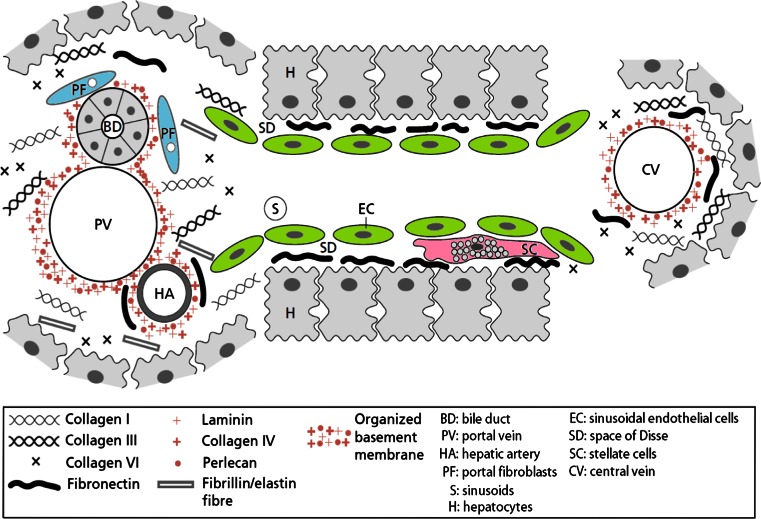
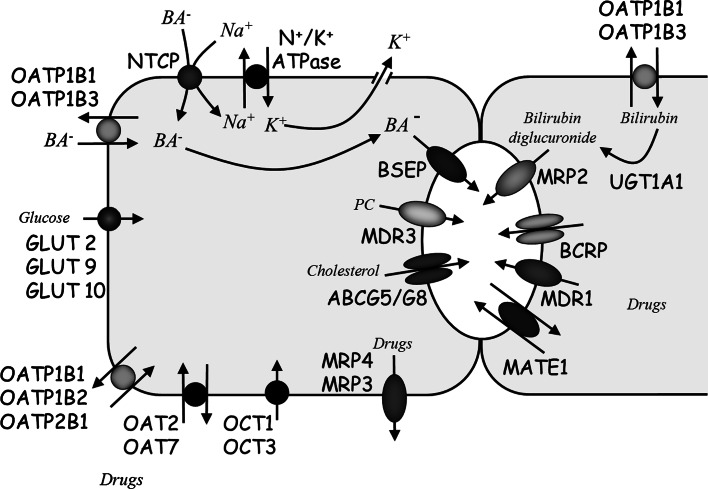


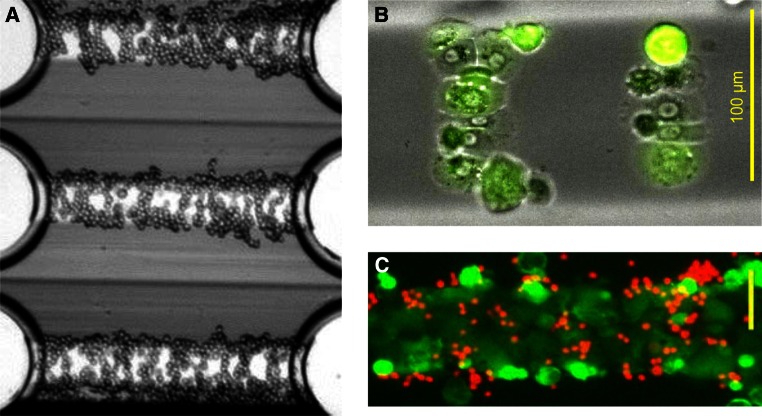
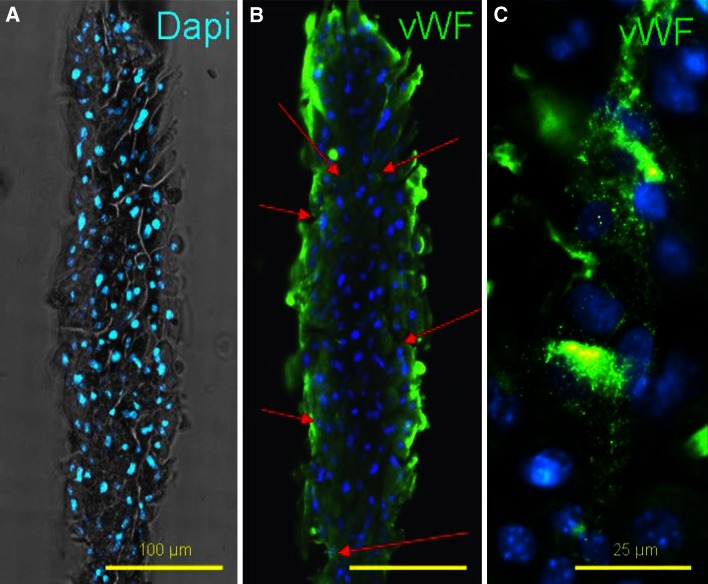
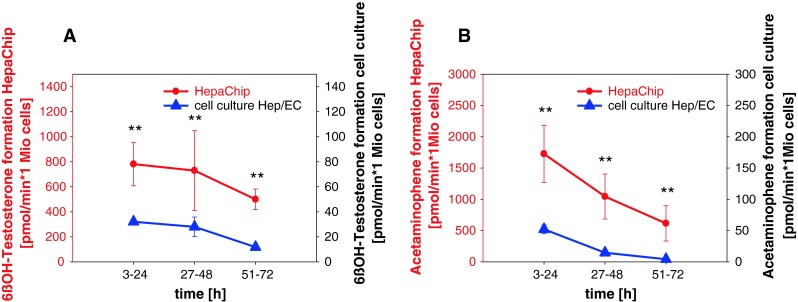
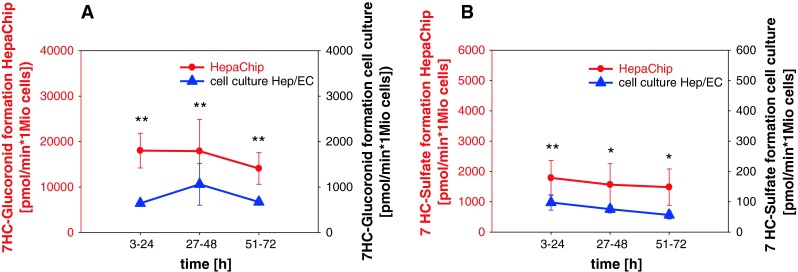
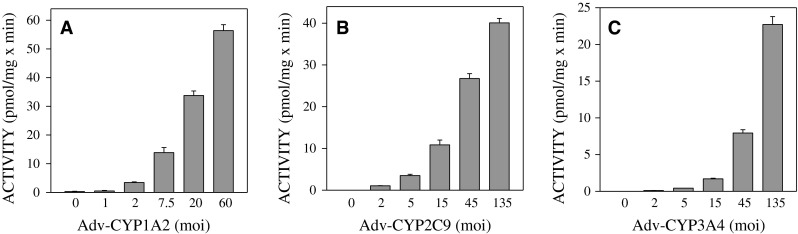
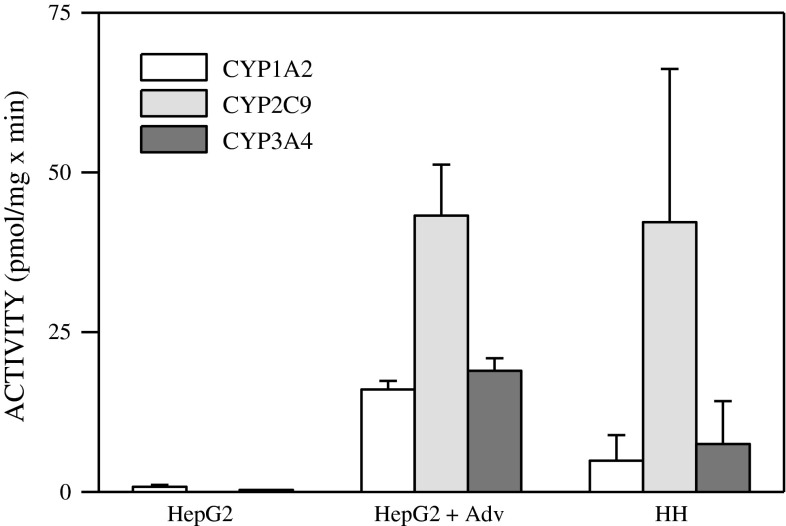
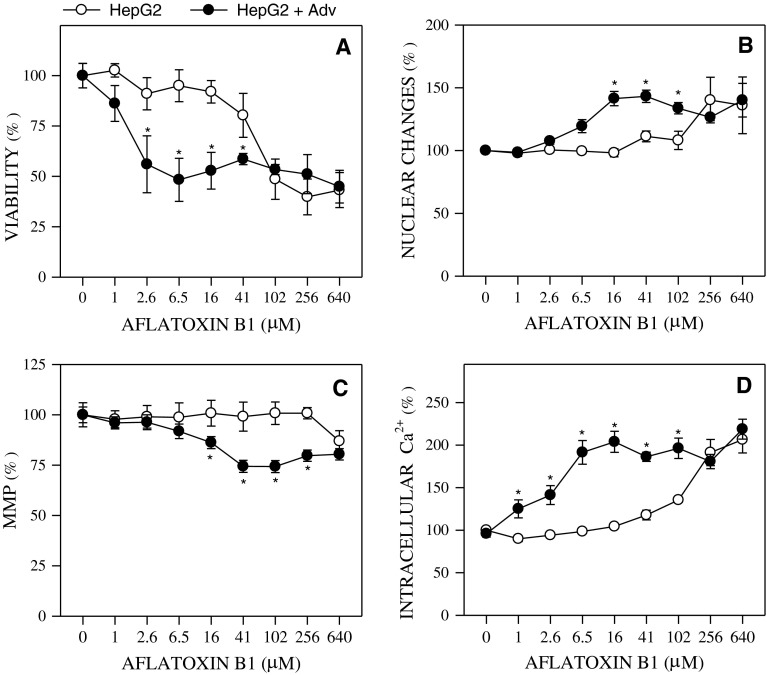
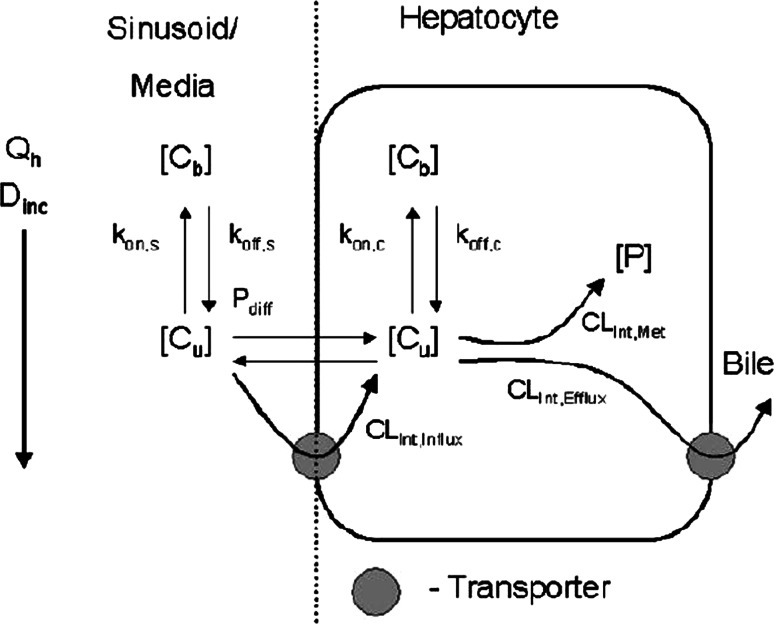




References
-
- Abe T, Unno M, Onogawa T, Tokui T, Kondo TN, Nakagomi R, Adachi H, Fujiwara K, Okabe M, Suzuki T, Nunoki K, Sato E, Kakyo M, Nishio T, Sugita J, Asano N, Tanemoto M, Seki M, Date F, Ono K, Kondo Y, Shiiba K, Suzuki M, Ohtani H, Shimosegawa T, Iinuma K, Nagura H, Ito S, Matsuno S. LST-2, a human liver-specific organic anion transporter, determines methotrexate sensitivity in gastrointestinal cancers. Gastroenterology. 2001;120:1689–1699. - PubMed
-
- Abramovitch S, Dahan-Bachar L, Sharvit E, Weisman Y, Bentov A, Brazowski E, Reif S. Vitamin D inhibits proliferation and profibrotic marker expression in hepatic stellate cells and decreases thioacetamide-induced liver fibrosis in rats. Gut. 2011;60:1728–37. - PubMed
-
- Abshagen K, Eipel C, Kalff JC, Menger MD, Vollmar B. Loss of NF-kappaB activation in Kupffer cell-depleted mice impairs liver regeneration after partial hepatectomy. Am J Physiol Gastrointest Liver Physiol. 2007;292:G1570–7. - PubMed
-
- Abshagen K, Eipel C, Kalff JC, Menger MD, Vollmar B. Kupffer cells are mandatory for adequate liver regeneration by mediating hyperperfusion via modulation of vasoactive proteins. Microcirculation. 2008;15:37–47. - PubMed
-
- Abu-Absi SF, Friend JR, Hansen LK, Hu WS. Structural polarity and functional bile canaliculi in rat hepatocyte spheroids. Exp Cell Res. 2002;274:56–67. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
