

Resveratrol accelerates erythroid maturation by activation of FoxO3 and ameliorates anemia in beta-thalassemic mice
- PMID: 23975182
- PMCID: PMC3912956
- DOI: 10.3324/haematol.2013.090076
Resveratrol accelerates erythroid maturation by activation of FoxO3 and ameliorates anemia in beta-thalassemic mice
Abstract
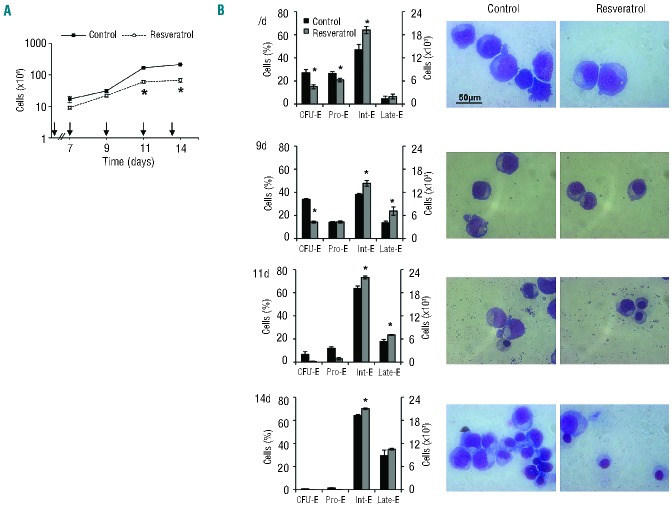
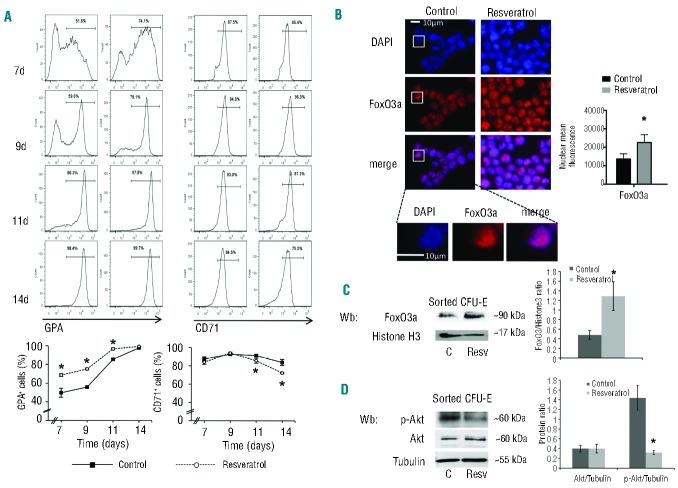
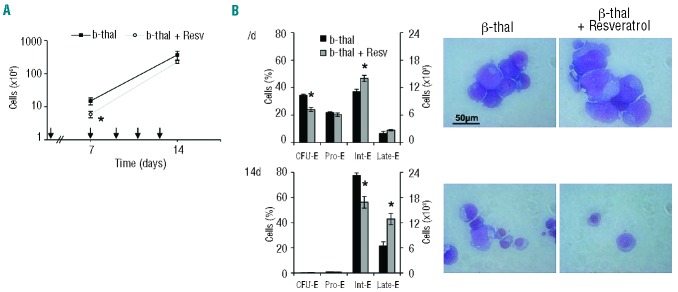
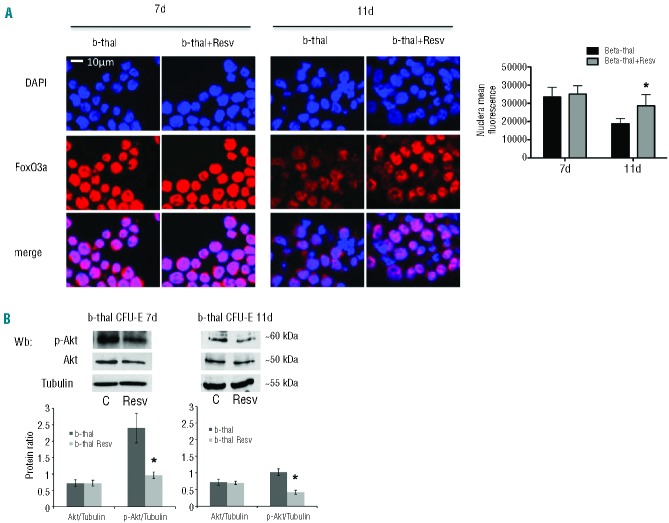
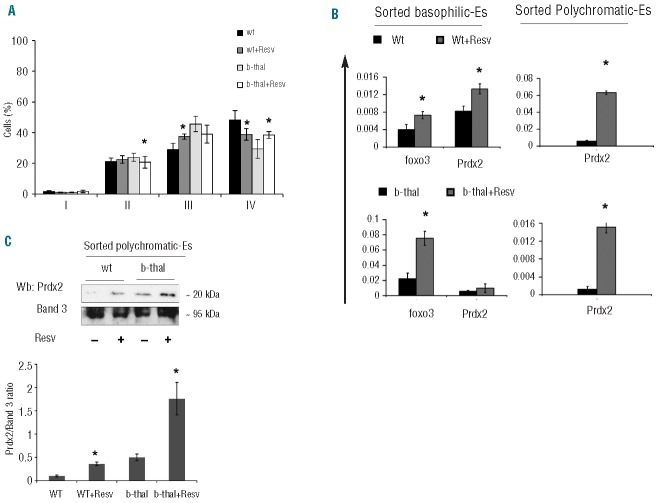
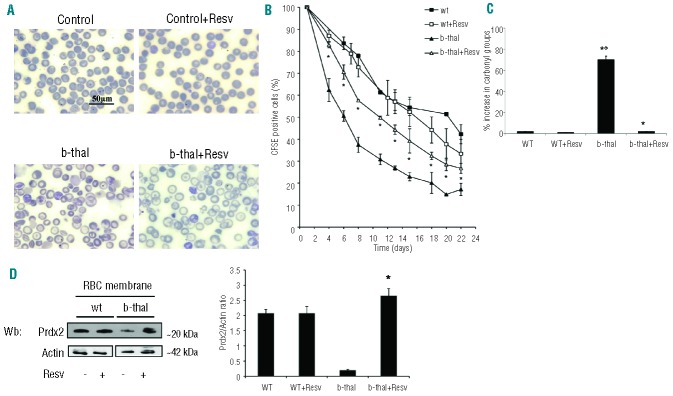
Resveratrol, a polyphenolic-stilbene, has received increased attention in the last decade due to its wide range of biological activities. Beta(β)-thalassemias are inherited red cell disorders, found worldwide, characterized by ineffective erythropoiesis and red cell oxidative damage with reduced survival. We evaluated the effects of low-dose-resveratrol (5 μM) on in vitro human erythroid differentiation of CD34(+) from normal and β-thalassemic subjects. We found that resveratrol induces accelerated erythroid-maturation, resulting in the reduction of colony-forming units of erythroid cells and increased intermediate and late erythroblasts. In sorted colony-forming units of erythroid cells resveratrol activates Forkhead-box-class-O3, decreases Akt activity and up-regulates anti-oxidant enzymes as catalase. In an in vivo murine model for β-thalassemia, resveratrol (2.4 mg/kg) reduces ineffective erythropoiesis, increases hemoglobin levels, reduces reticulocyte count and ameliorates red cell survival. In both wild-type and β-thalassemic mice, resveratrol up-regulates scavenging enzymes such as catalase and peroxiredoxin-2 through Forkhead-box-class-O3 activation. These data indicate that resveratrol inhibits Akt resulting in FoxO3 activation with upregulation of cytoprotective systems enabling the pathological erythroid precursors to resist the oxidative damage and continue to differentiate. Our data suggest that the dual effect of resveratrol on erythropoiesis through activation of FoxO3 transcriptional factor combined with the amelioration of oxidative stress in circulating red cells may be considered as a potential novel therapeutic strategy in treating β-thalassemia.
Figures
References
-
- Cucciolla V, Borriello A, Oliva A, Galletti P, Zappia V, Della Ragione F. Resveratrol: from basic science to the clinic. Cell Cycle. 2007; 6(20):2495–510 - PubMed
-
- Ferry-Dumazet H, Garnier O, Mamani-Matsuda M, Vercauteren J, Belloc F, Billiard C, et al. Resveratrol inhibits the growth and induces the apoptosis of both normal and leukemic hematopoietic cells. Carcinogenesis. 2002;23(8):1327–33 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
