

An Integrative Review of Mechanotransduction in Endothelial, Epithelial (Renal) and Dendritic Cells (Osteocytes)
- PMID: 23976901
- PMCID: PMC3748935
- DOI: 10.1007/s12195-011-0179-6
An Integrative Review of Mechanotransduction in Endothelial, Epithelial (Renal) and Dendritic Cells (Osteocytes)
Abstract
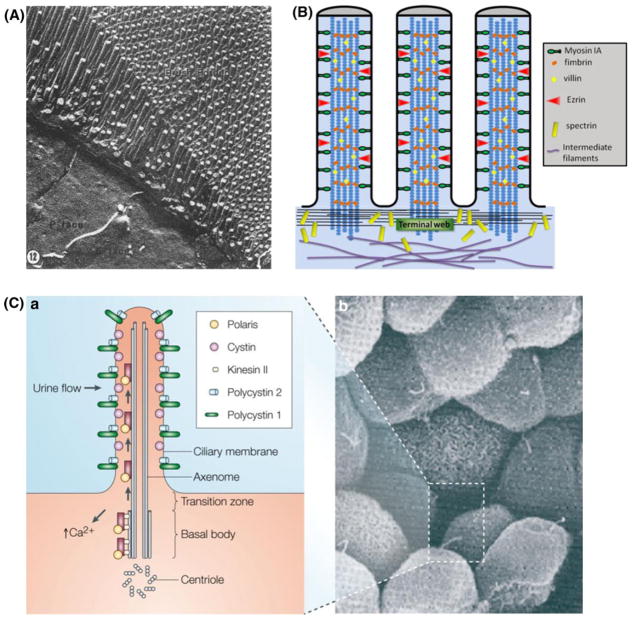
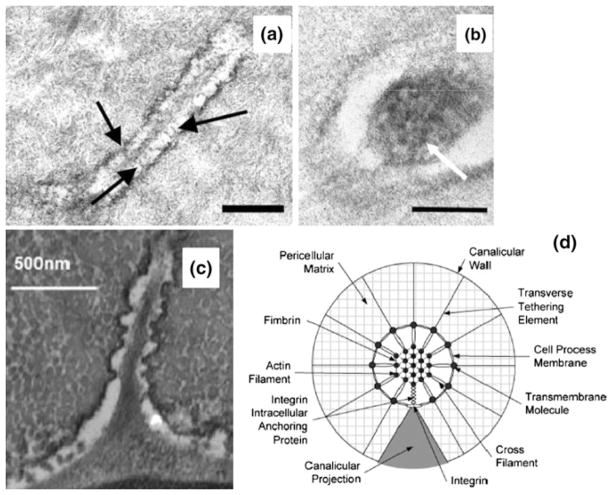
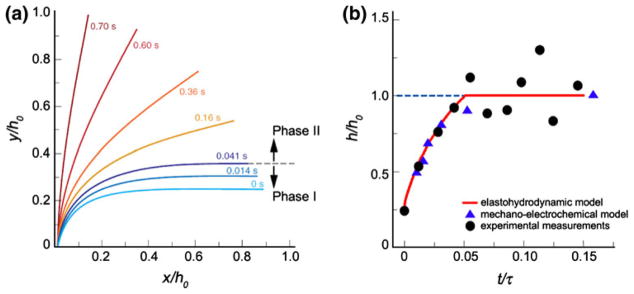
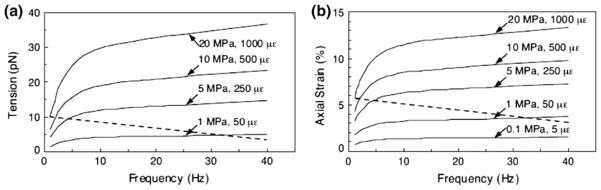
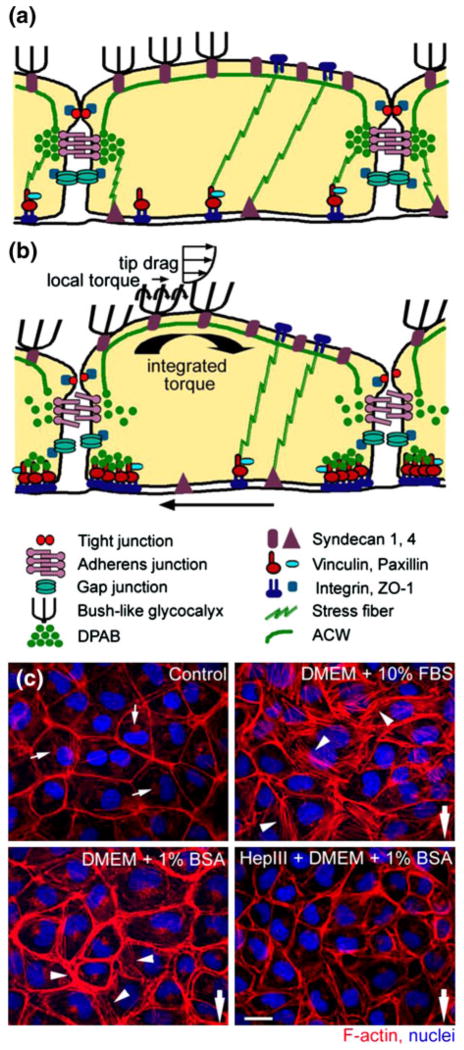
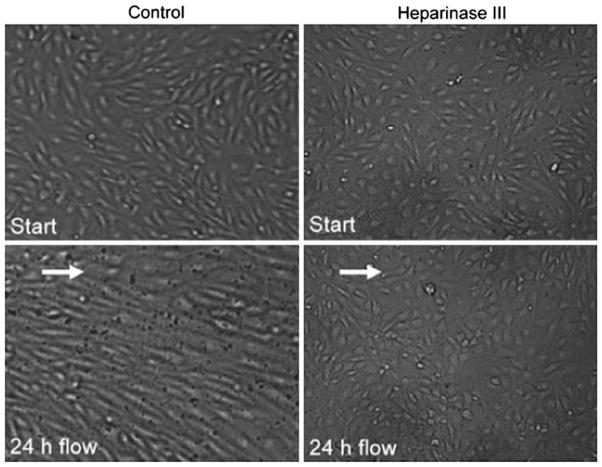
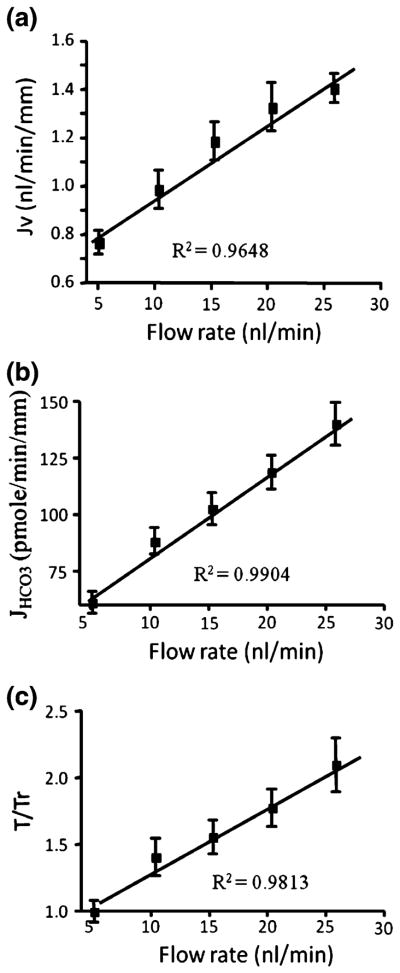
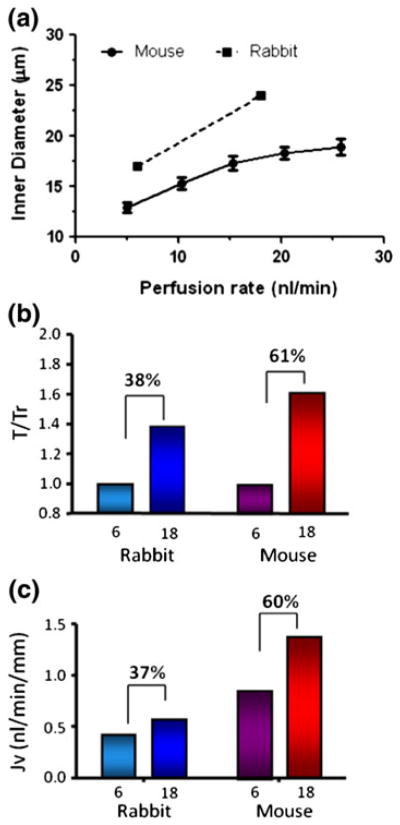
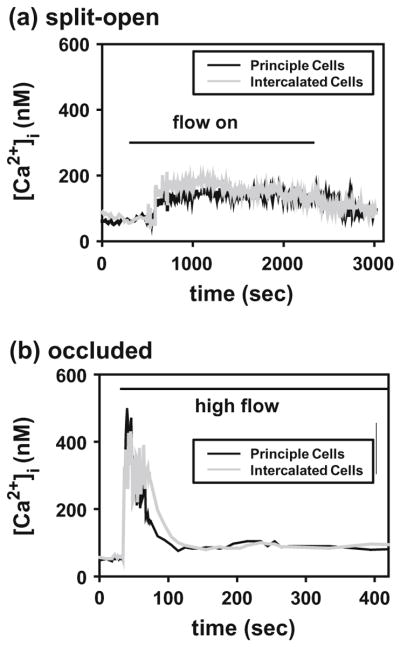
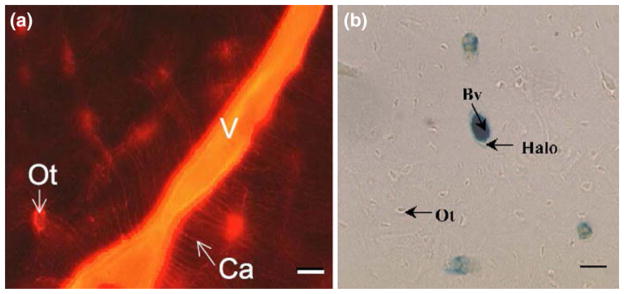
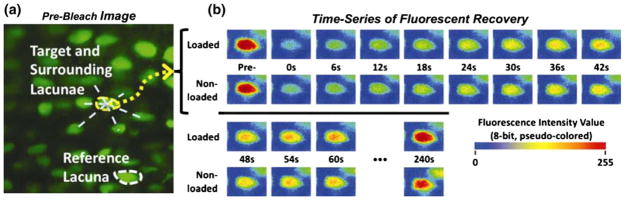
In this review we will examine from a biomechanical and ultrastructural viewpoint how the cytoskeletal specialization of three basic cell types, endothelial cells (ECs), epithelial cells (renal tubule) and dendritic cells (osteocytes), enables the mechano-sensing of fluid flow in both their native in vivo environment and in culture, and the downstream signaling that is initiated at the molecular level in response to fluid flow. These cellular responses will be discussed in terms of basic mysteries and paradoxes encountered by each cell type. In ECs fluid shear stress (FSS) is nearly entirely attenuated by the endothelial glycocalyx that covers their apical membrane and yet FSS is communicated to both intracellular and junctional molecular components in activating a wide variety of signaling pathways. The same is true in proximal tubule (PT) cells where a dense brush border of microvilli covers the apical surface and the flow at the apical membrane is negligible. A four decade old unexplained mystery is the ability of PT epithelia to reliably reabsorb 60% of the flow entering the tubule regardless of the glomerular filtration rate. In the cortical collecting duct (CCD) the flow rates are so low that a special sensing apparatus, a primary cilia is needed to detect very small variations in tubular flow. In bone it has been a century old mystery as to how osteocytes embedded in a stiff mineralized tissue are able to sense miniscule whole tissue strains that are far smaller than the cellular level strains required to activate osteocytes in vitro.
Keywords: Actin cortical web; Actin filament bundles; Bone cell processes; Brush border microvilli; Cortical collecting duct; Endothelial glycocalyx; Integrin attachments; Lacunar-canalicular system; Proximal tubule.
Figures

References
-
- Adachi T, Aonuma Y, Tanaka M, Hojo M, Takano-Yamamoto T, Kamioka H. Calcium response in single osteocytes to locally applied mechanical stimulus: differences in cell process and cell body. J Biomech. 2009;42:1989–1995. - PubMed
-
- Akst J. Full speed ahead: physical forces acting in and around cells are fast—and making waves in the world of molecular biology. Scientist. 2009;23:26–32.
-
- Alenghat FJ, Nauli SM, Kolb R, Zhou J, Ingber DE. Global cytoskeletal control of mechanotransduction in kidney epithelial cells. Exp Cell Res. 2004;301:23–30. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
