

On the measurement of ecological novelty: scale-eating pupfish are separated by 168 my from other scale-eating fishes
- PMID: 23976994
- PMCID: PMC3747246
- DOI: 10.1371/journal.pone.0071164
On the measurement of ecological novelty: scale-eating pupfish are separated by 168 my from other scale-eating fishes
Abstract
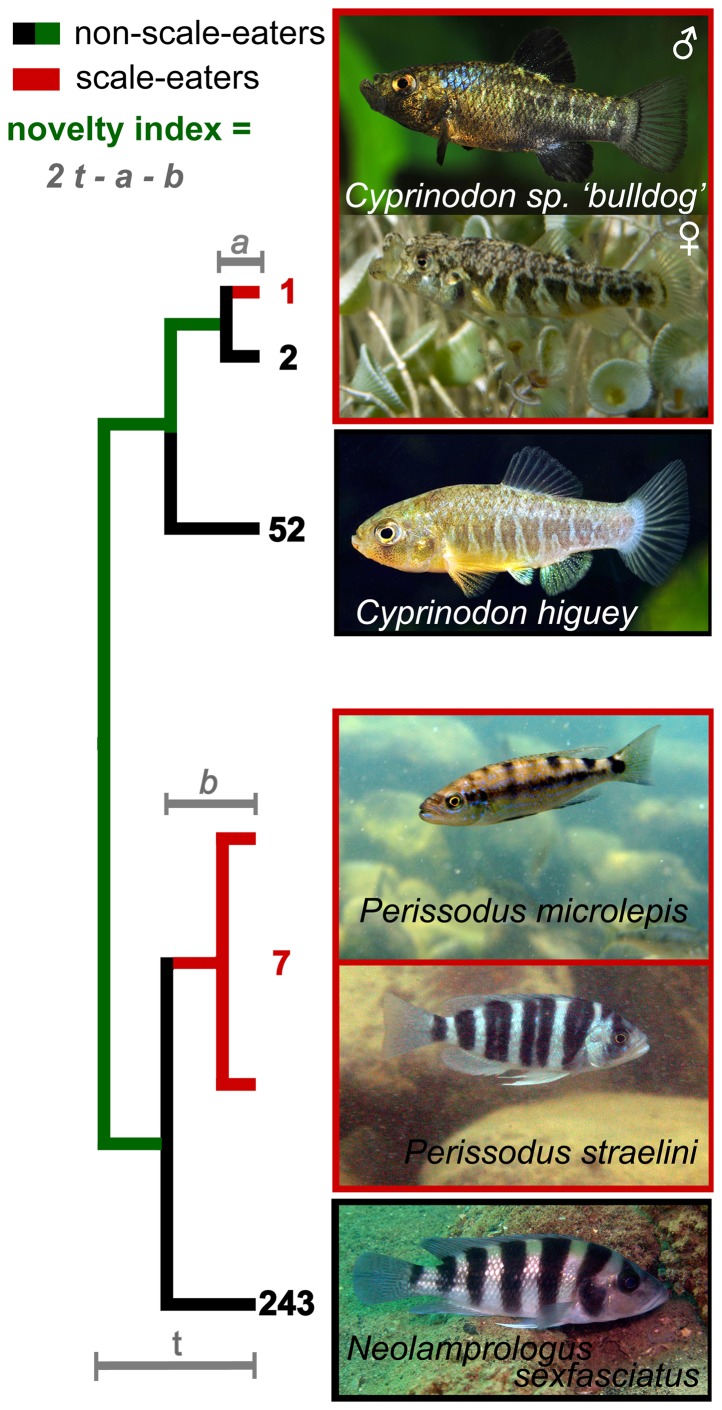
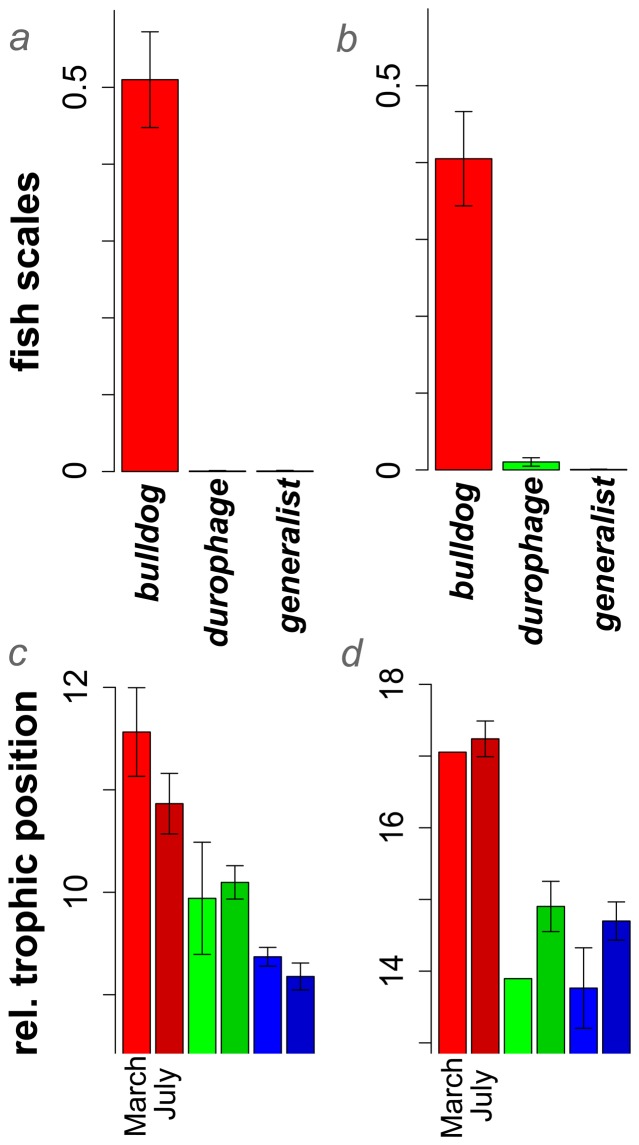
The colonization of new adaptive zones is widely recognized as one of the hallmarks of adaptive radiation. However, the adoption of novel resources during this process is rarely distinguished from phenotypic change because morphology is a common proxy for ecology. How can we quantify ecological novelty independent of phenotype? Our study is split into two parts: we first document a remarkable example of ecological novelty, scale-eating (lepidophagy), within a rapidly-evolving adaptive radiation of Cyprinodon pupfishes on San Salvador Island, Bahamas. This specialized predatory niche is known in several other fish groups, but is not found elsewhere among the 1,500 species of atherinomorphs. Second, we quantify this ecological novelty by measuring the time-calibrated phylogenetic distance in years to the most closely-related species with convergent ecology. We find that scale-eating pupfish are separated by 168 million years of evolution from the nearest scale-eating fish. We apply this approach to a variety of examples and highlight the frequent decoupling of ecological novelty from phenotypic divergence. We observe that novel ecology is not always tightly correlated with rates of phenotypic or species diversification, particularly within recent adaptive radiations, necessitating the use of additional measures of ecological novelty independent of phenotype.
Conflict of interest statement
Figures
References
-
- Moczek AP (2008) On the origins of novelty in development and evolution. BioEssays 30: 432–47. - PubMed
-
- Lefebvre L, Reader SM, Sol D (2004) Brains, innovations and evolution in birds and primates. Brain Behav Evol 63: 233–46. - PubMed
-
- Muller GB, Wagner GP (1991) Novelty in evolution: restructuring the concept. Ann Rev Ecol Syst 22: 229–256.
-
- Nitecki NH (1990) Evolutionary Innovations. University of Chicago Press, Chicago.
-
- Givnish TJ, Sytsma KJ, Smith JF, Hahn WJ, Benzing DH, et al. (1997) Molecular evolution and adaptive radiation in Brocchinia (Bromeliaceae: Pitcairnioideae) atop tepuis of the Guayana Shield. In: Molecular evolution and adaptive radiation (Givnish, TJ, Sytsma, KJ, eds), Cambridge University Press, Cambridge.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
