

More than just a cargo adapter, melanophilin prolongs and slows processive runs of myosin Va
- PMID: 23979131
- PMCID: PMC3795233
- DOI: 10.1074/jbc.M113.476929
More than just a cargo adapter, melanophilin prolongs and slows processive runs of myosin Va
Abstract
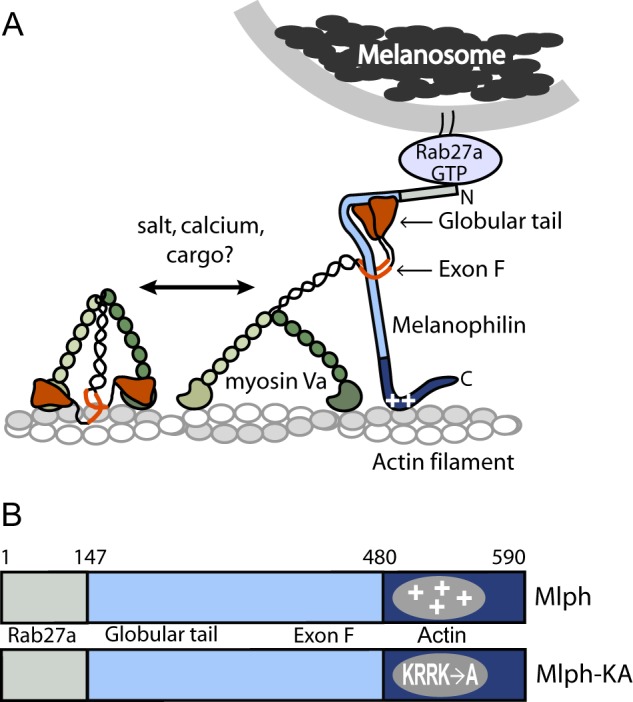
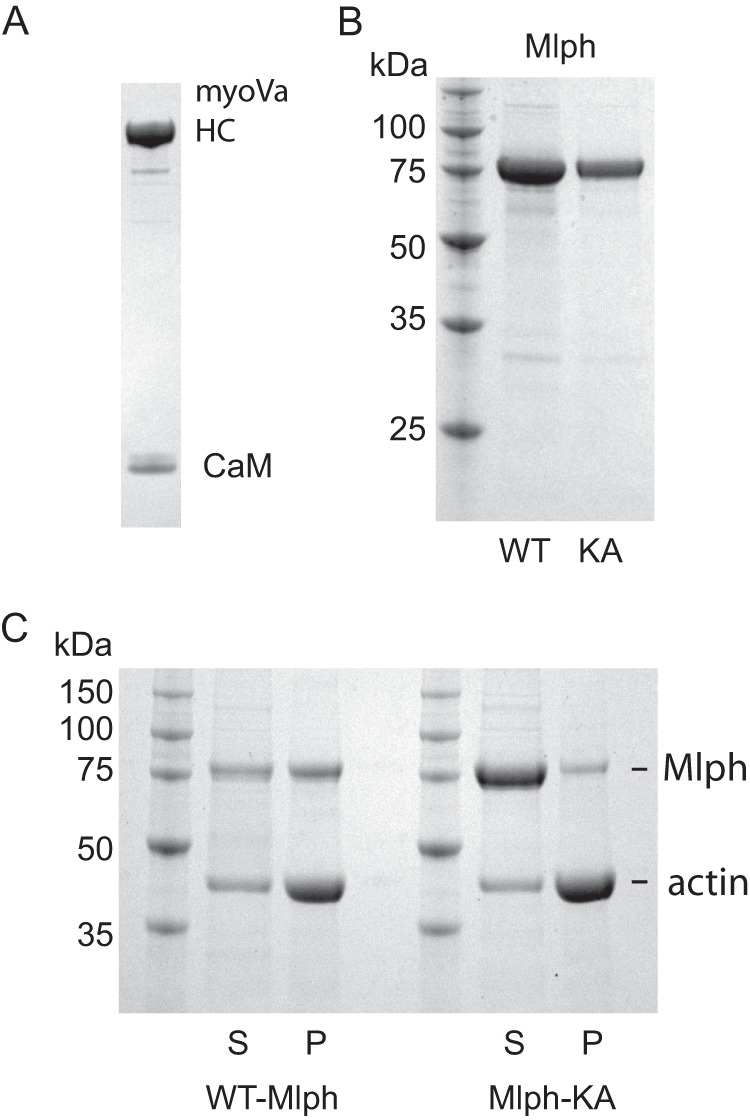
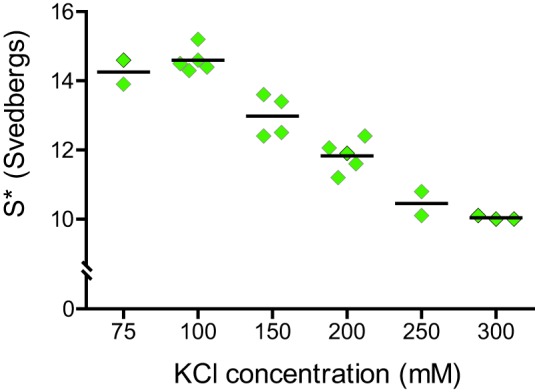
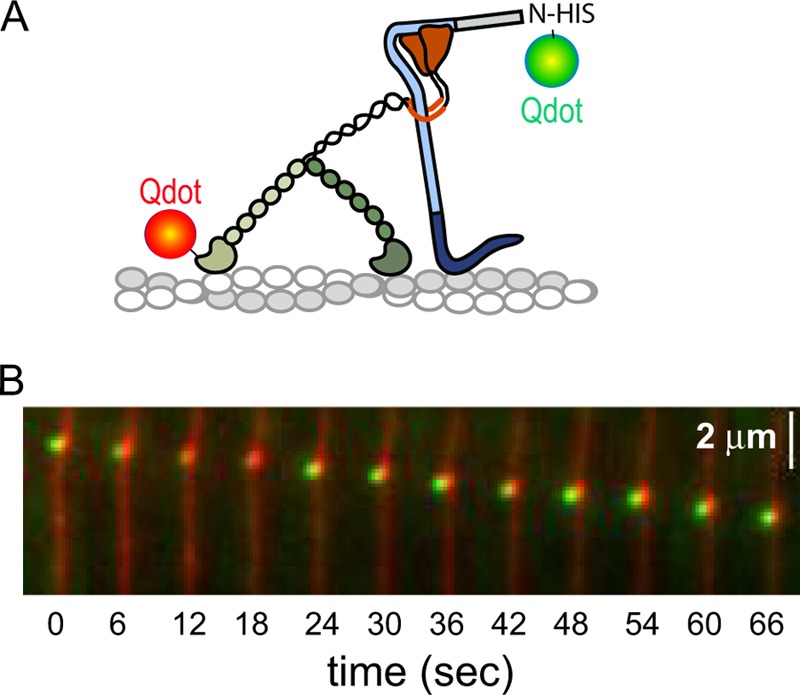
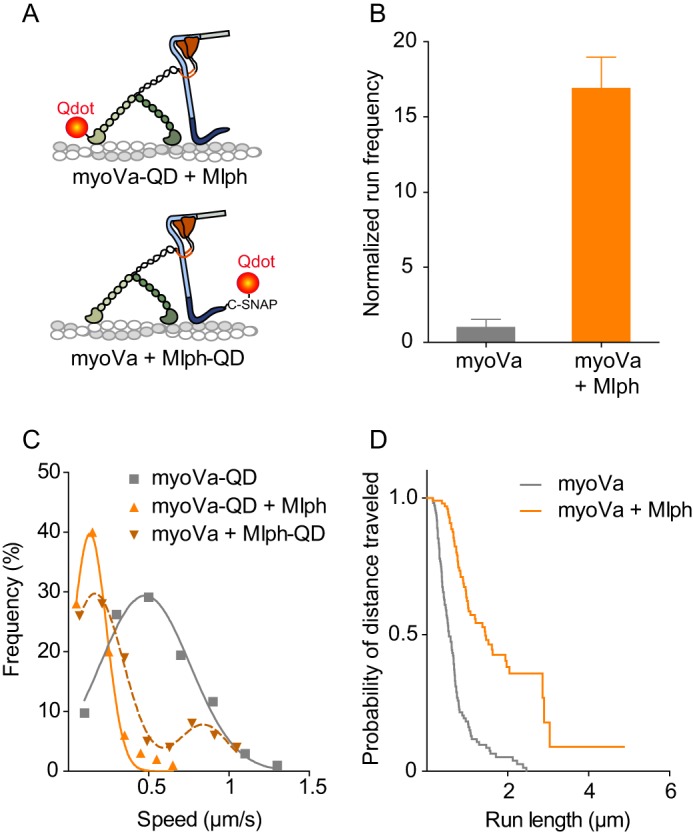
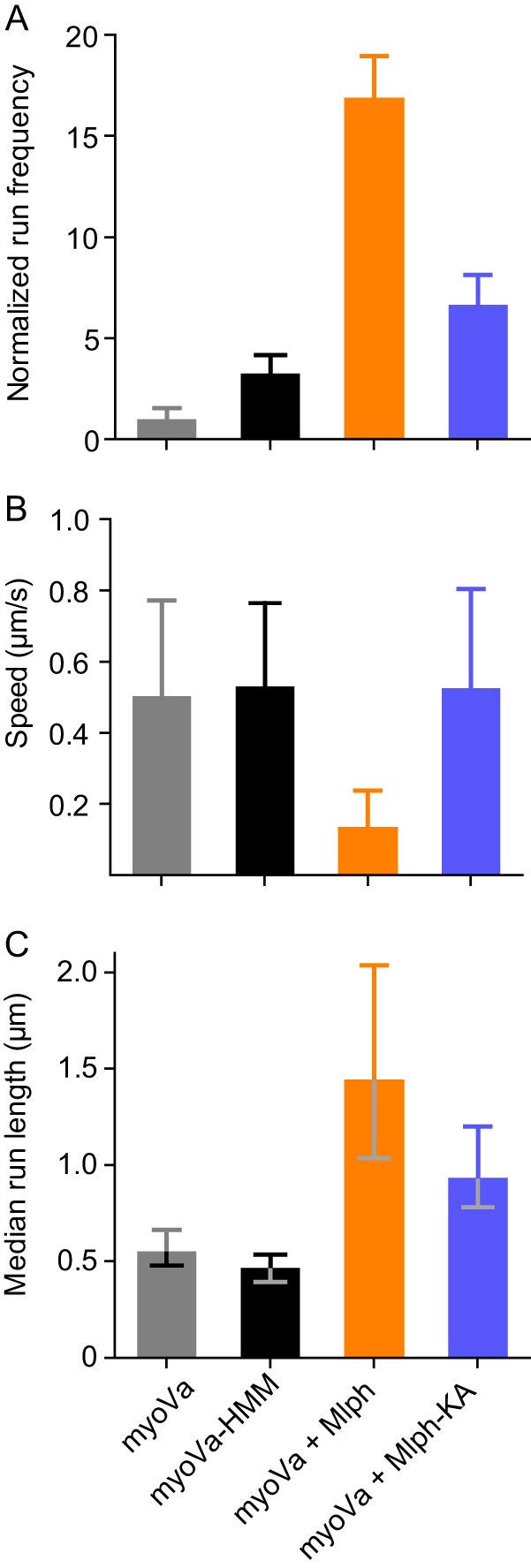
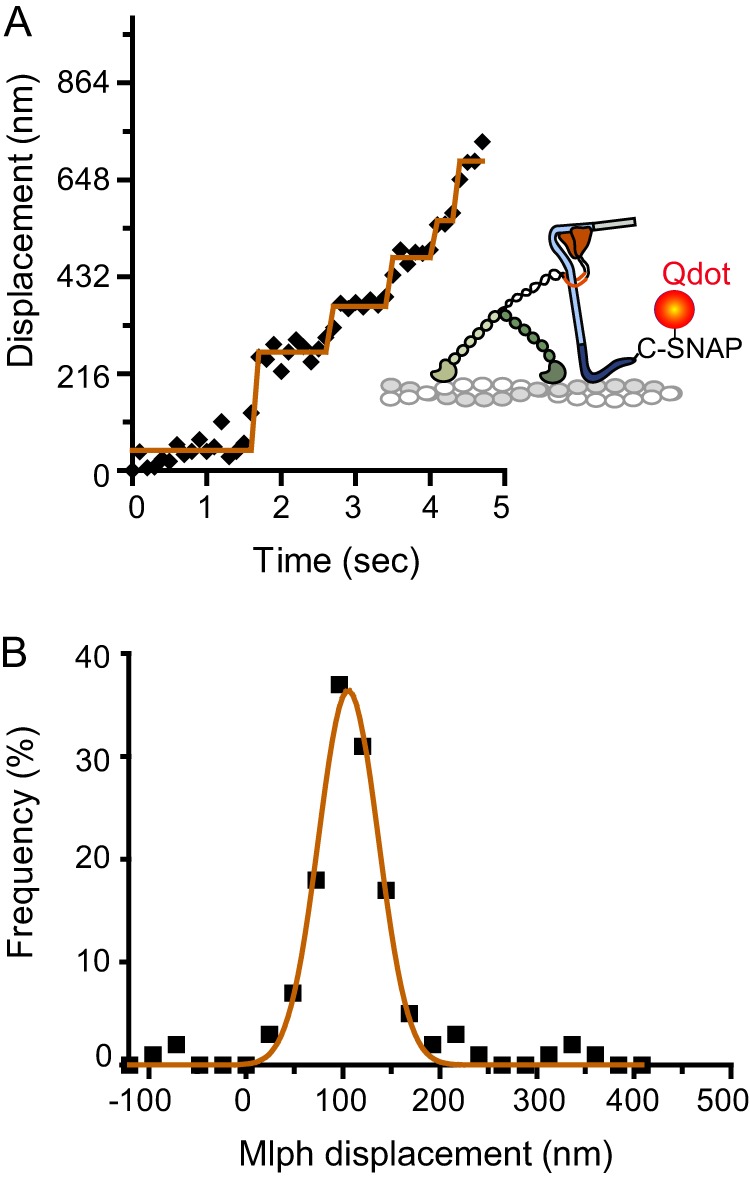
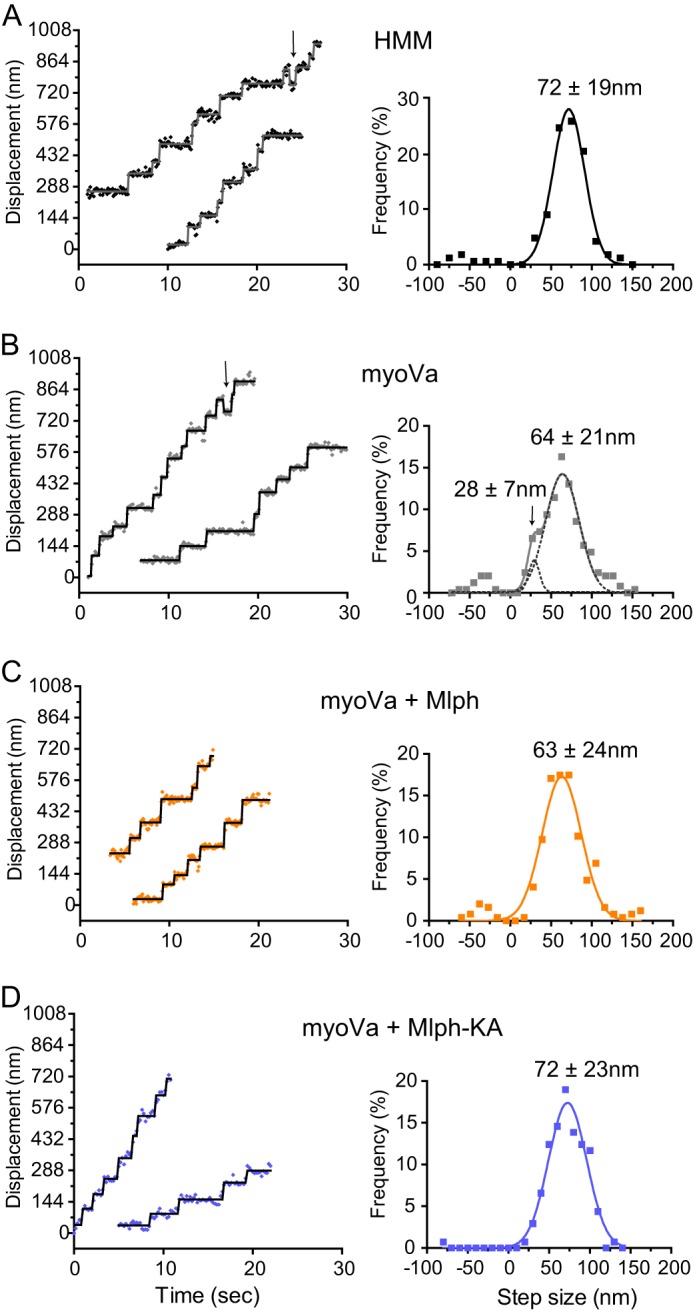
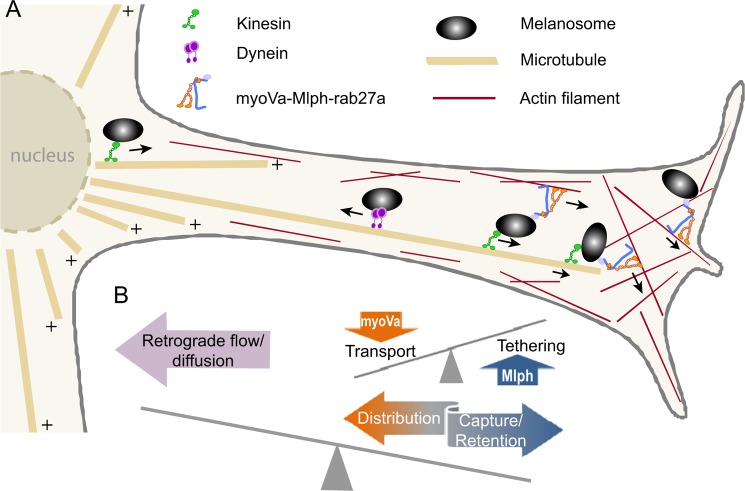
Myosin Va (myoVa) is a molecular motor that processively transports cargo along actin tracks. One well studied cargo in vivo is the melanosome, a pigment organelle that is moved first by kinesin on microtubules and then handed off to myoVa for transport in the actin-rich dendritic periphery of melanocytes. Melanophilin (Mlph) is the adapter protein that links Rab27a-melanosomes to myoVa. Using total internal reflection fluorescence microscopy and quantum dot-labeled full-length myoVa, we show at the single-molecule level that Mlph increases the number of processively moving myoVa motors by 17-fold. Surprisingly, myoVa-Mlph moves ~4-fold slower than myoVa alone and with twice the run length. These two changes greatly increase the time spent on actin, a property likely to enhance the transfer of melanosomes to the adjacent keratinocyte. In contrast to the variable stepping pattern of full-length myoVa, the myoVa-Mlph complex shows a normal gating pattern between the heads typical of a fully active motor and consistent with a cargo-dependent activation mechanism. The Mlph-dependent changes in myoVa depend on a positively charged cluster of amino acids in the actin binding domain of Mlph, suggesting that Mlph acts as a "tether" that links the motor to the track. Our results provide a molecular explanation for the uncharacteristically slow speed of melanosome movement by myoVa in vivo. More generally, these data show that proteins that link motors to cargo can modify motor properties to enhance their biological role.
Keywords: Gating; Intracellular Trafficking; Melanophilin; Myosin; Nanotechnology; Processivity; Protein Dynamics; Regulation; Slac-2a; Tethering.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
