

The duration of sleep promoting efficacy by dual orexin receptor antagonists is dependent upon receptor occupancy threshold
- PMID: 23981345
- PMCID: PMC3765993
- DOI: 10.1186/1471-2202-14-90
The duration of sleep promoting efficacy by dual orexin receptor antagonists is dependent upon receptor occupancy threshold
Abstract
Background: Drugs targeting insomnia ideally promote sleep throughout the night, maintain normal sleep architecture, and are devoid of residual effects associated with morning sedation. These features of an ideal compound are not only dependent upon pharmacokinetics, receptor binding kinetics, potency and pharmacodynamic activity, but also upon a compound's mechanism of action.
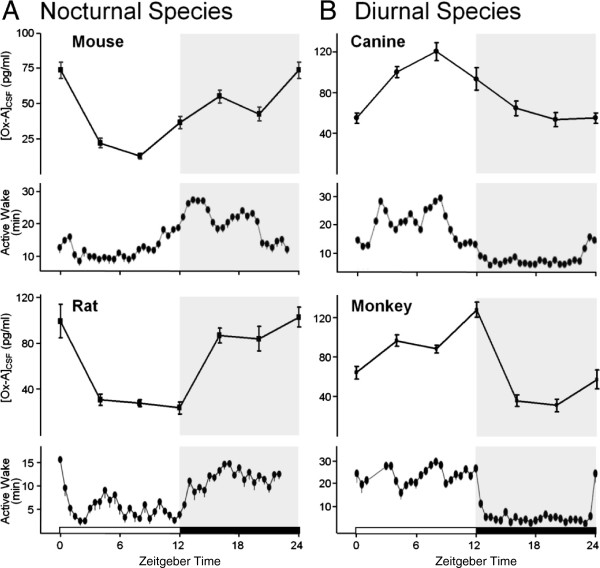
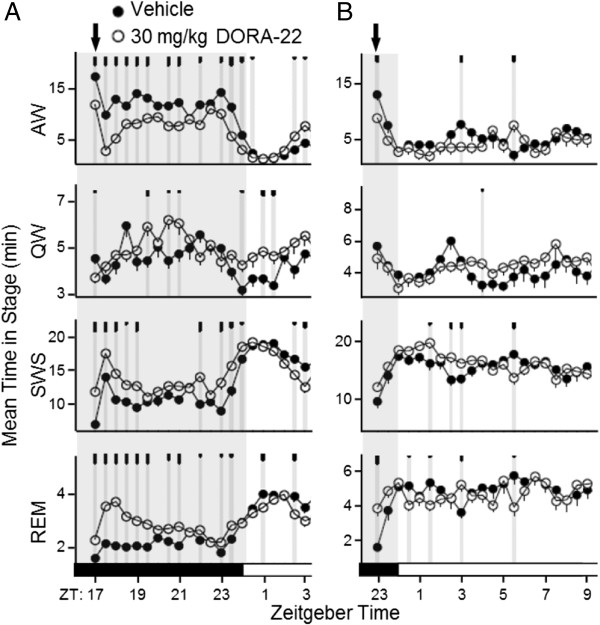
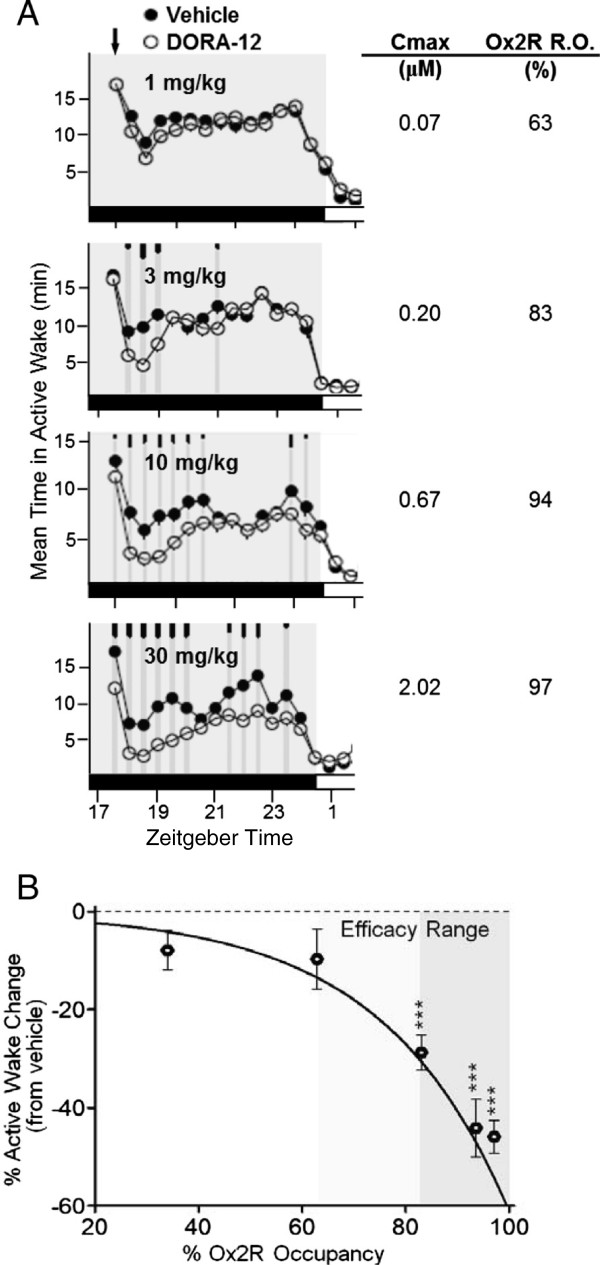
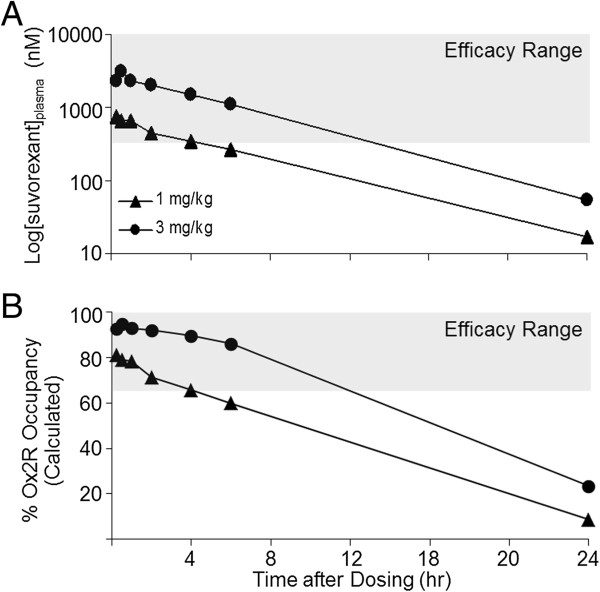
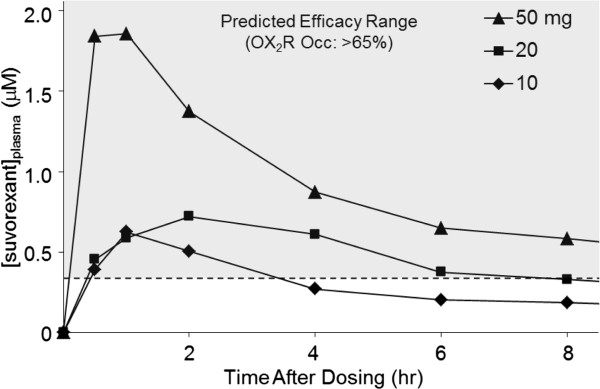
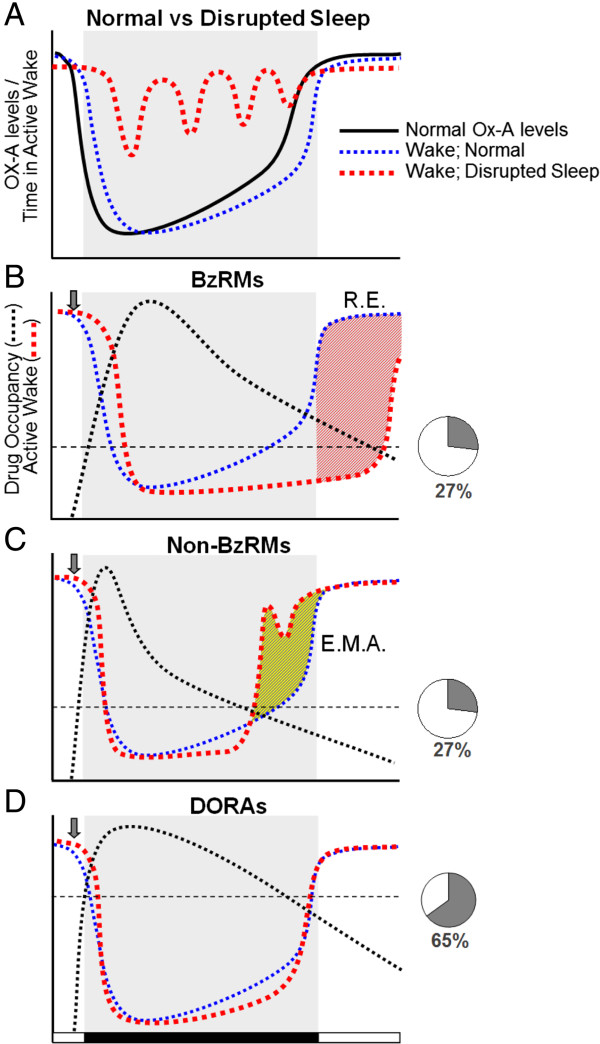
Results: Dual orexin receptor antagonists (DORAs) block the arousal-promoting activity of orexin peptides and, as demonstrated in the current work, exhibit an efficacy signal window dependent upon oscillating levels of endogenous orexin neuropeptide. Sleep efficacy of structurally diverse DORAs in rat and dog was achieved at plasma exposures corresponding to orexin 2 receptor (OX2R) occupancies in the range of 65 to 80%. In rats, the time course of OX2R occupancy was dependent upon receptor binding kinetics and was tightly correlated with the timing of active wake reduction. In rhesus monkeys, direct comparison of DORA-22 with GABA-A modulators at similar sleep-inducing doses revealed that diazepam produced next-day residual sleep and both diazepam and eszopiclone induced next-day cognitive deficits. In stark contrast, DORA-22 did not produce residual effects. Furthermore, DORA-22 evoked only minimal changes in quantitative electroencephalogram (qEEG) activity during the normal resting phase in contrast to GABA-A modulators which induced substantial qEEG changes.
Conclusion: The higher levels of receptor occupancy necessary for DORA efficacy require a plasma concentration profile sufficient to maintain sleep for the duration of the resting period. DORAs, with a half-life exceeding 8 h in humans, are expected to fulfill this requirement as exposures drop to sub-threshold receptor occupancy levels prior to the wake period, potentially avoiding next-day residual effects at therapeutic doses.
Figures


References
-
- De Lecea L, Kilduff TS, Peyron C, Gao XB, Foye PE, Danielson PE, Fukuhara C, Battenberg ELF, Gautvik VT, Bartlett FS, Frankel WN, van den Pol AN, Bloom FE, Gautvik KM, Sutcliffe JG. The hypocretins: Hypothalamus-specific peptides with neuroexcitatory activity. Proc Natl Acad Sci U S A. 1998;95:322–327. doi: 10.1073/pnas.95.1.322. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
