

How the unstable eye sees a stable and moving world
- PMID: 23988388
- PMCID: PMC4525302
- DOI: 10.1167/13.10.22
How the unstable eye sees a stable and moving world
Abstract
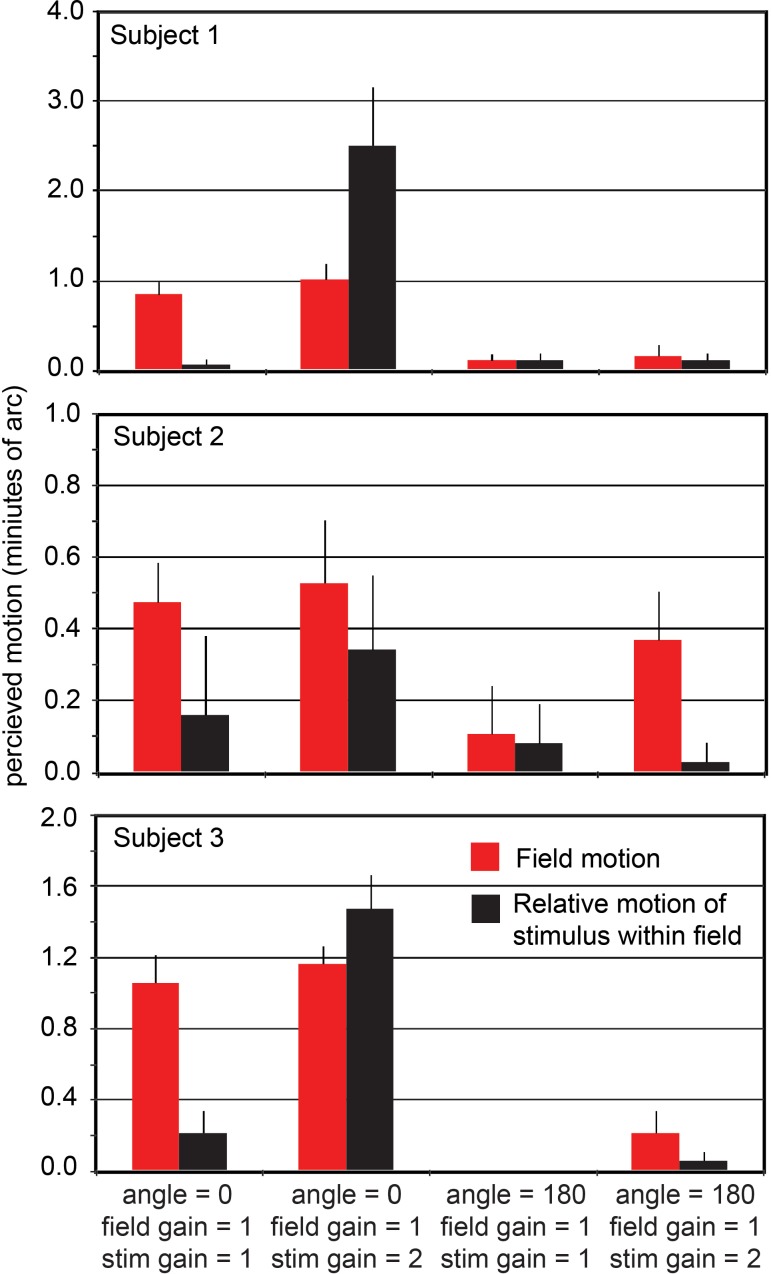
Eye motion, even during fixation, results in constant motion of the image of the world on our retinas. Vision scientists have long sought to understand the process by which we perceive the stable parts of the world as unmoving despite this instability and perceive the moving parts with realistic motion. We used an instrument capable of delivering visual stimuli with controlled motion relative to the retina at cone-level precision while capturing the subjects' percepts of stimulus motion with a matching task. We found that the percept of stimulus motion is more complex than conventionally thought. Retinal stimuli that move in a direction that is consistent with eye motion (i.e., opposite eye motion) appear stable even if the magnitude of that motion is amplified. The apparent stabilization diminishes for stimulus motions increasingly inconsistent with eye motion direction. Remarkably, we found that this perceived direction-contingent stabilization occurs separately for each separately moving pattern on the retina rather than for the image as a whole. One consequence is that multiple patterns that move at different rates relative to each other in the visual input are perceived as immobile with respect to each other, thereby disrupting our hyperacute sensitivity to target motion against a frame of reference. This illusion of relative stability has profound implications regarding the underlying visual mechanisms. Functionally, the system compensates retinal slip induced by eye motion without requiring an extremely precise optomotor signal and, at the same time, retains an exquisite sensitivity to an object's true motion in the world.
Keywords: adaptive optics; eye tracking; motion perception.
Figures
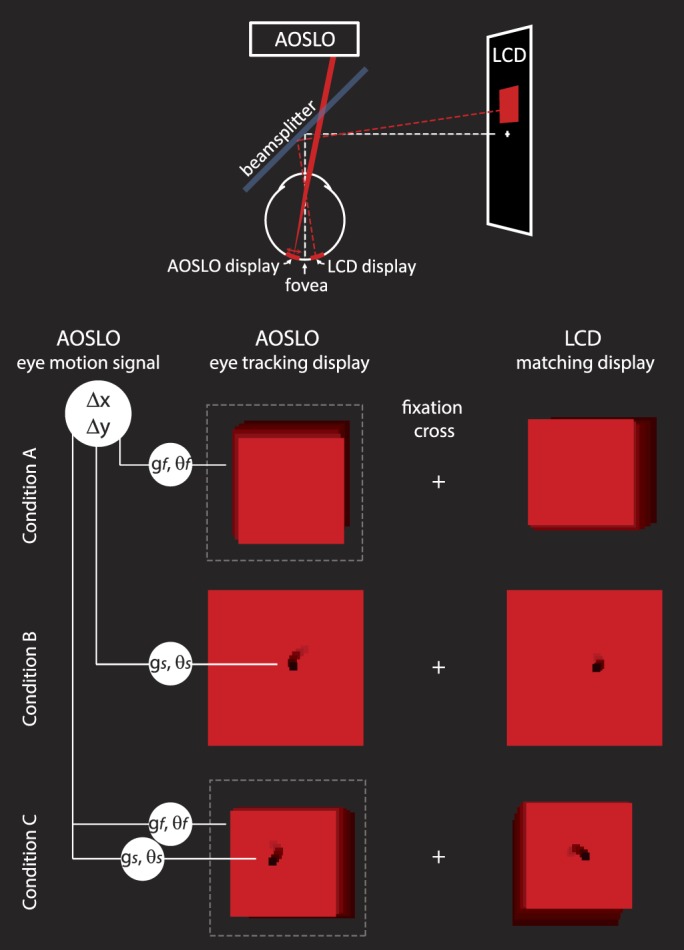

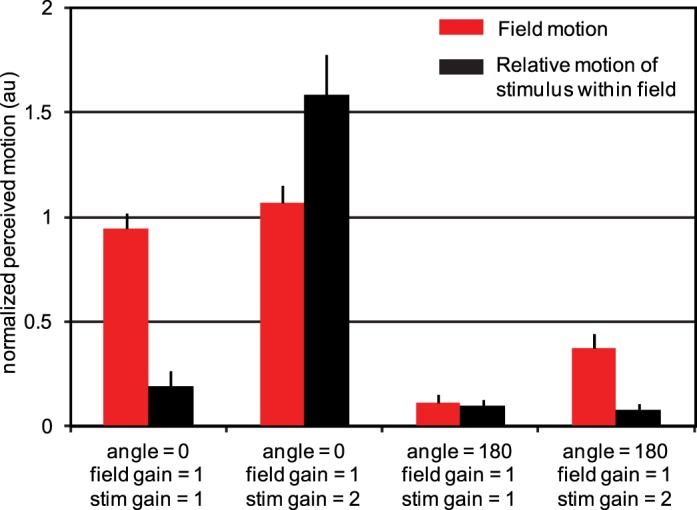
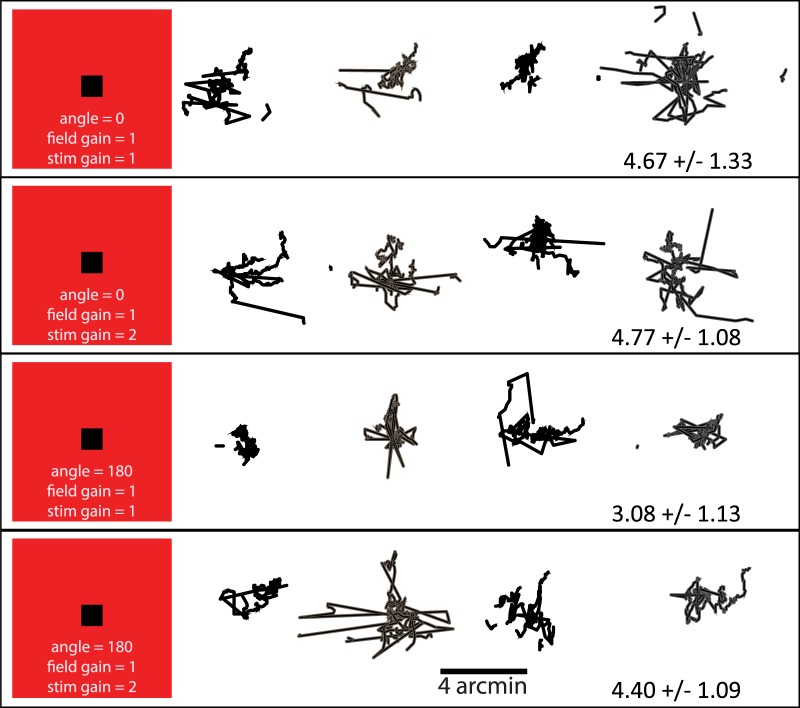
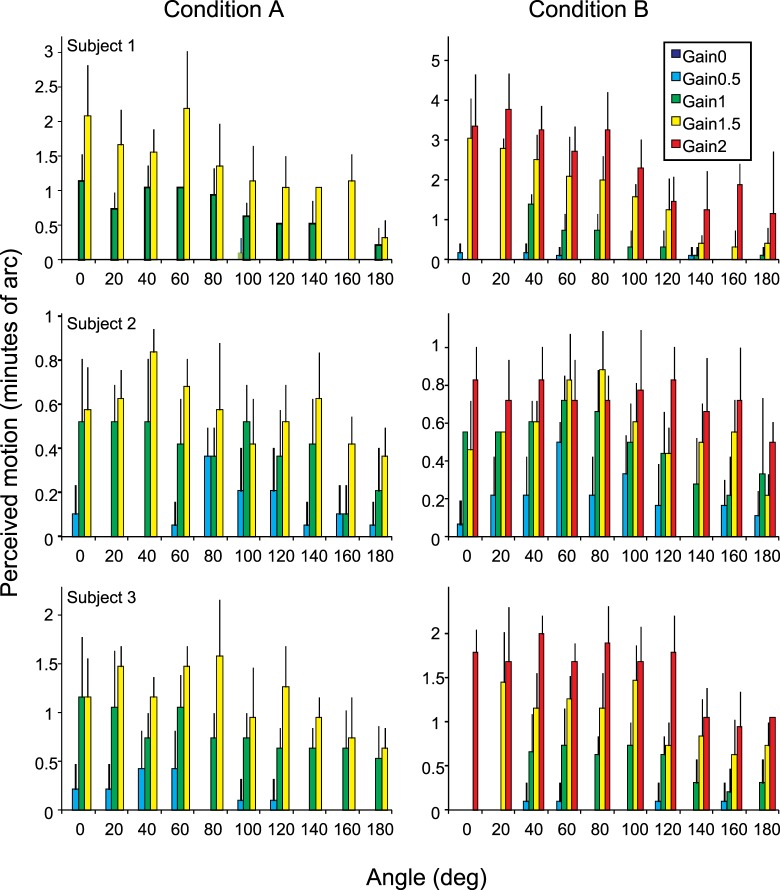
References
-
- Arathorn D. W. (2002). Map-seeking circuits in visual cognition. Stanford: Stanford University Press.
-
- Arathorn D. W., Yang Q., Vogel C. R., Zhang Y., Tiruveedhula P., Roorda A. (2007). Retinally stabilized cone-targeted stimulus delivery. Optics Express , 15, 13731–13744. - PubMed
-
- Bridgeman B., Hendry D., Stark L. (1975). Failure to detect displacement of the visual world during saccadic eye movements. Vision Research , 15 (6), 719–722. - PubMed
-
- Bridgeman B., Stark L. (1991). Ocular proprioception and efference copy in registering visual direction. Vision Research , 31 (11), 1903–1913. - PubMed
-
- Cornsweet T. N. (1956). Determination of the stimuli for involuntary drifts and saccadic eye movements. Journal of the Optical Society of America , 46 (11), 987–993. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
