

PPARα and Sirt1 mediate erythropoietin action in increasing metabolic activity and browning of white adipocytes to protect against obesity and metabolic disorders
- PMID: 23990359
- PMCID: PMC3837041
- DOI: 10.2337/db13-0518
PPARα and Sirt1 mediate erythropoietin action in increasing metabolic activity and browning of white adipocytes to protect against obesity and metabolic disorders
Abstract
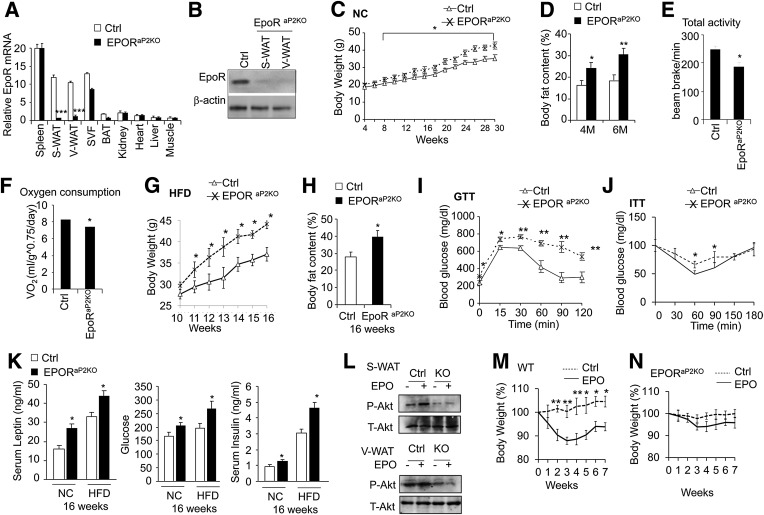
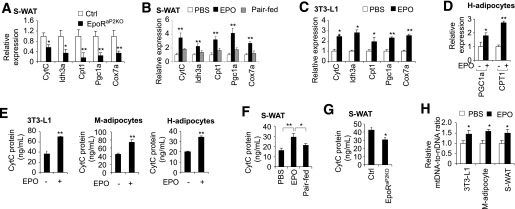
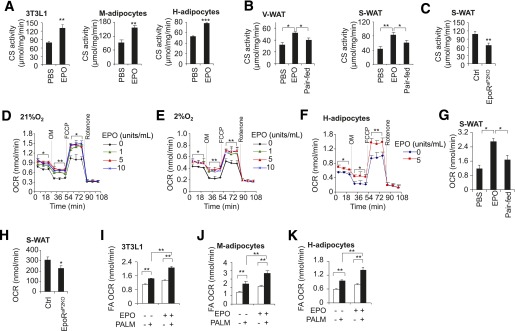
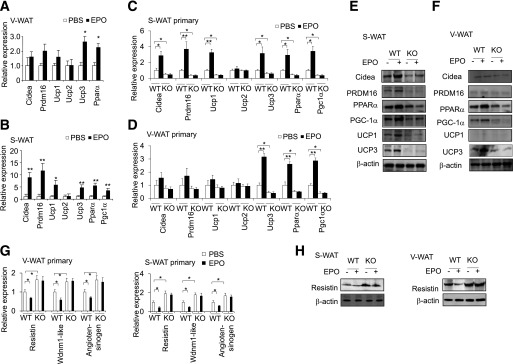
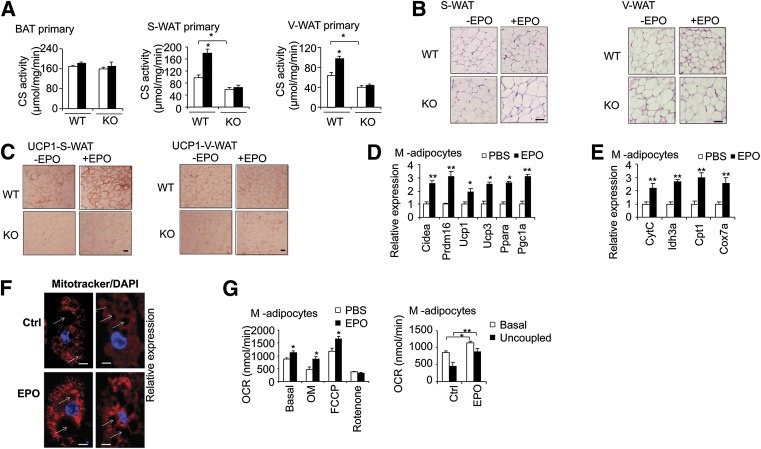
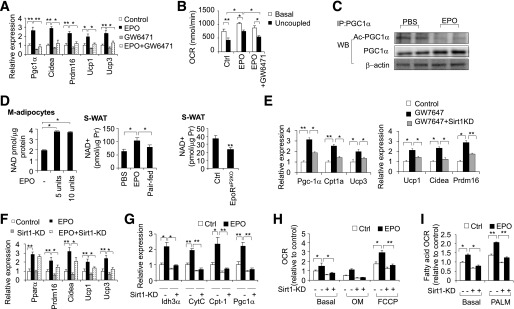
Erythropoietin (EPO) has shown beneficial effects in the regulation of obesity and metabolic syndrome; however, the detailed mechanism is still largely unknown. Here, we created mice with adipocyte-specific deletion of EPO receptor. These mice exhibited obesity and decreased glucose tolerance and insulin sensitivity, especially when fed a high-fat diet. Moreover, EPO increased oxidative metabolism, fatty acid oxidation, and key metabolic genes in adipocytes and in white adipose tissue from diet-induced obese wild-type mice. Increased metabolic activity by EPO is associated with induction of brown fat-like features in white adipocytes, as demonstrated by increases in brown fat gene expression, mitochondrial content, and uncoupled respiration. Peroxisome proliferator-activated receptor (PPAR)α was found to mediate EPO activity because a PPARα antagonist impaired EPO-mediated induction of brown fat-like gene expression and uncoupled respiration. PPARα also cooperates with Sirt1 activated by EPO through modulating the NAD+ level to regulate metabolic activity. PPARα targets, including PPARγ coactivator 1α, uncoupling protein 1, and carnitine palmitoyltransferase 1α, were increased by EPO but impaired by Sirt1 knockdown. Sirt1 knockdown also attenuated adipose response to EPO. Collectively, EPO, as a novel regulator of adipose energy homeostasis via these metabolism coregulators, provides a potential therapeutic strategy to protect against obesity and metabolic disorders.
Figures
References
-
- Burger D, Lei M, Geoghegan-Morphet N, Lu X, Xenocostas A, Feng Q. Erythropoietin protects cardiomyocytes from apoptosis via up-regulation of endothelial nitric oxide synthase. Cardiovasc Res 2006;72:51–59 - PubMed
-
- Beleslin-Cokic BB, Cokic VP, Yu X, Weksler BB, Schechter AN, Noguchi CT. Erythropoietin and hypoxia stimulate erythropoietin receptor and nitric oxide production by endothelial cells. Blood 2004;104:2073–2080 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
