

Crystal structure of MraY, an essential membrane enzyme for bacterial cell wall synthesis
- PMID: 23990562
- PMCID: PMC3906829
- DOI: 10.1126/science.1236501
Crystal structure of MraY, an essential membrane enzyme for bacterial cell wall synthesis
Abstract
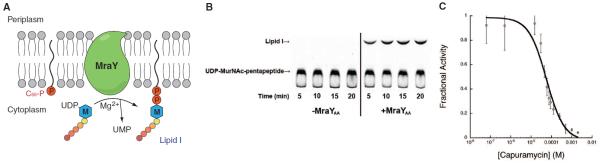
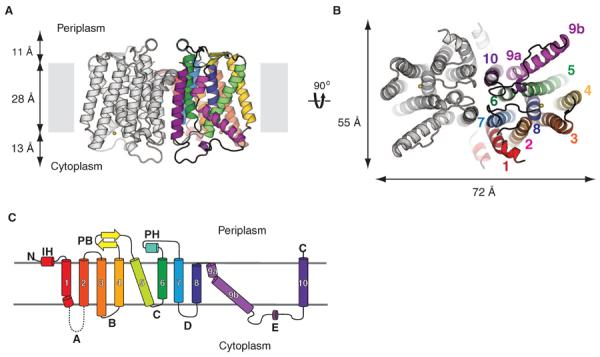
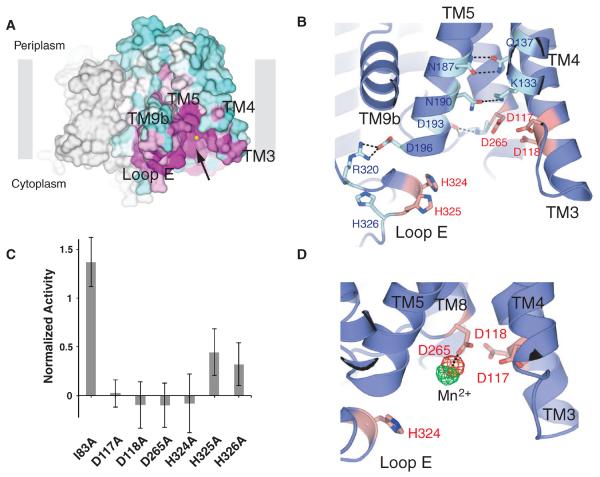
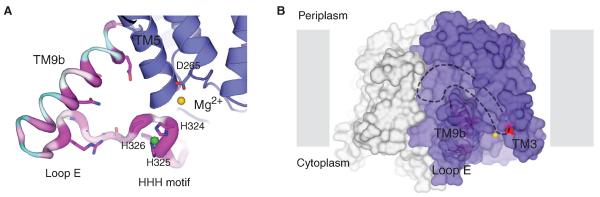
MraY (phospho-MurNAc-pentapeptide translocase) is an integral membrane enzyme that catalyzes an essential step of bacterial cell wall biosynthesis: the transfer of the peptidoglycan precursor phospho-MurNAc-pentapeptide to the lipid carrier undecaprenyl phosphate. MraY has long been considered a promising target for the development of antibiotics, but the lack of a structure has hindered mechanistic understanding of this critical enzyme and the enzyme superfamily in general. The superfamily includes enzymes involved in bacterial lipopolysaccharide/teichoic acid formation and eukaryotic N-linked glycosylation, modifications that are central in many biological processes. We present the crystal structure of MraY from Aquifex aeolicus (MraYAA) at 3.3 Å resolution, which allows us to visualize the overall architecture, locate Mg(2+) within the active site, and provide a structural basis of catalysis for this class of enzyme.
Figures
References
-
- Winn M, Goss RJ, Kimura K, Bugg TD. Nat. Prod. Rep. 2010;27:279–304. - PubMed
-
- Bugg TD, Braddick D, Dowson CG, Roper DI. Trends Biotechnol. 2011;29:167–173. - PubMed
-
- Bouhss A, Trunkfield AE, Bugg TD, Mengin-Lecreulx D. FEMS Microbiol. Rev. 2008;32:208–233. - PubMed
-
- Mendel S, Holbourn JM, Schouten JA, Bugg TD. Microbiology. 2006;152:2959–2967. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
