

The Merkel cell polyomavirus minor capsid protein
- PMID: 23990782
- PMCID: PMC3749969
- DOI: 10.1371/journal.ppat.1003558
The Merkel cell polyomavirus minor capsid protein
Abstract
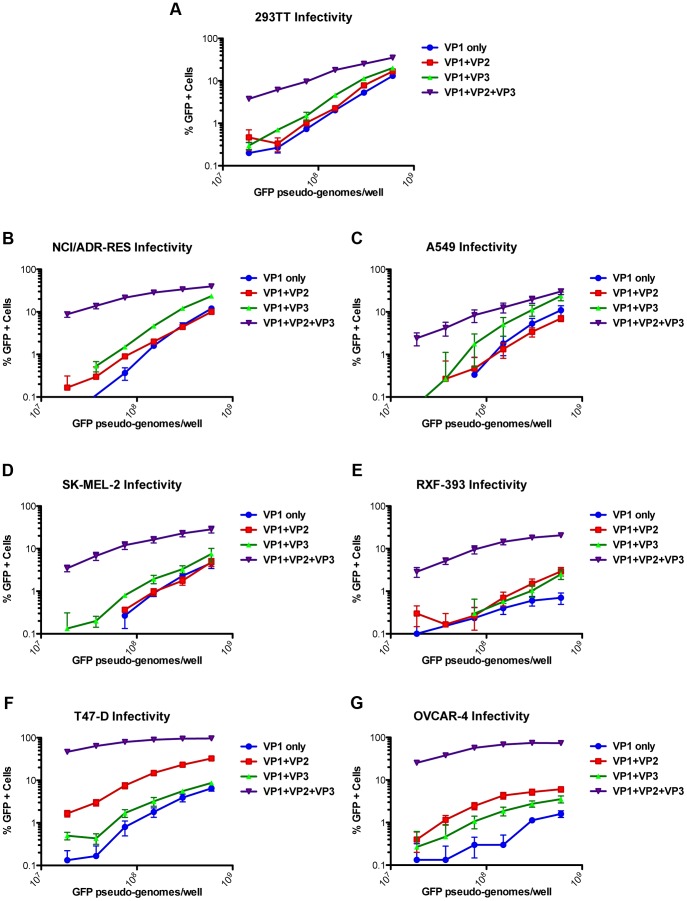
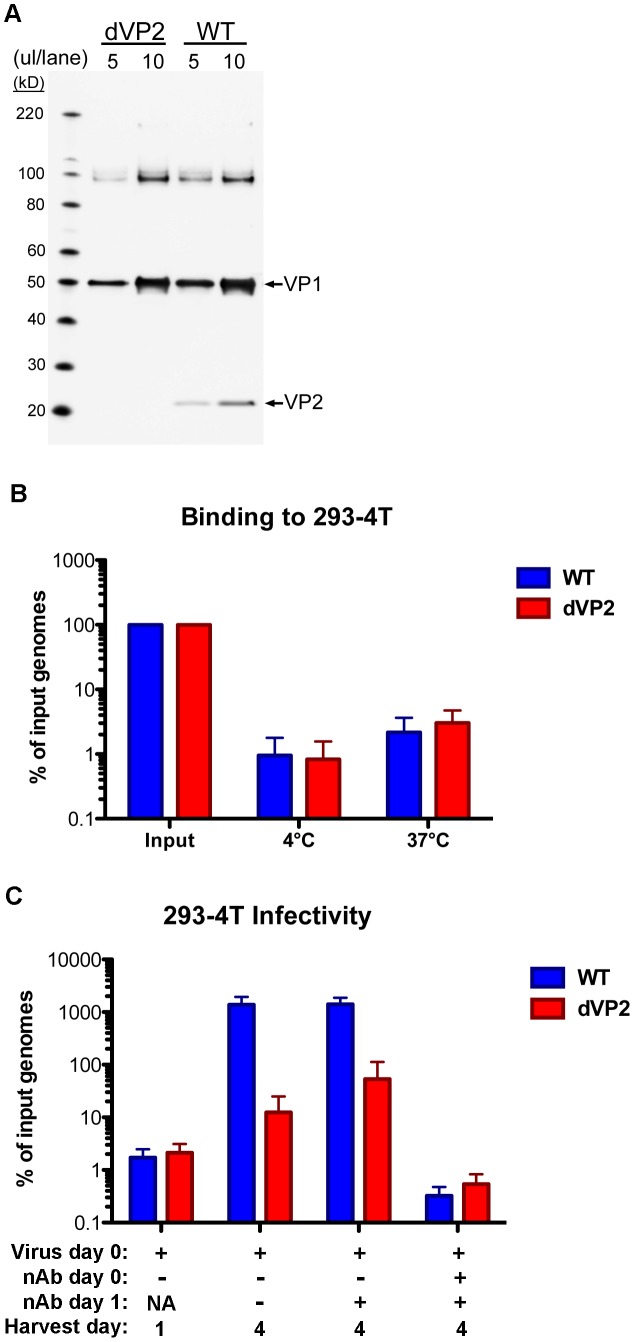
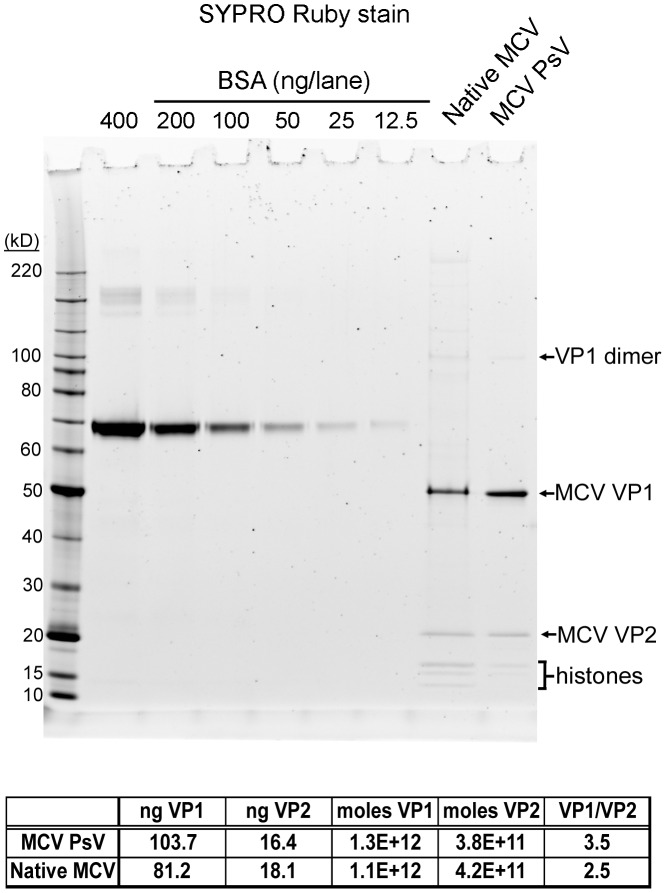
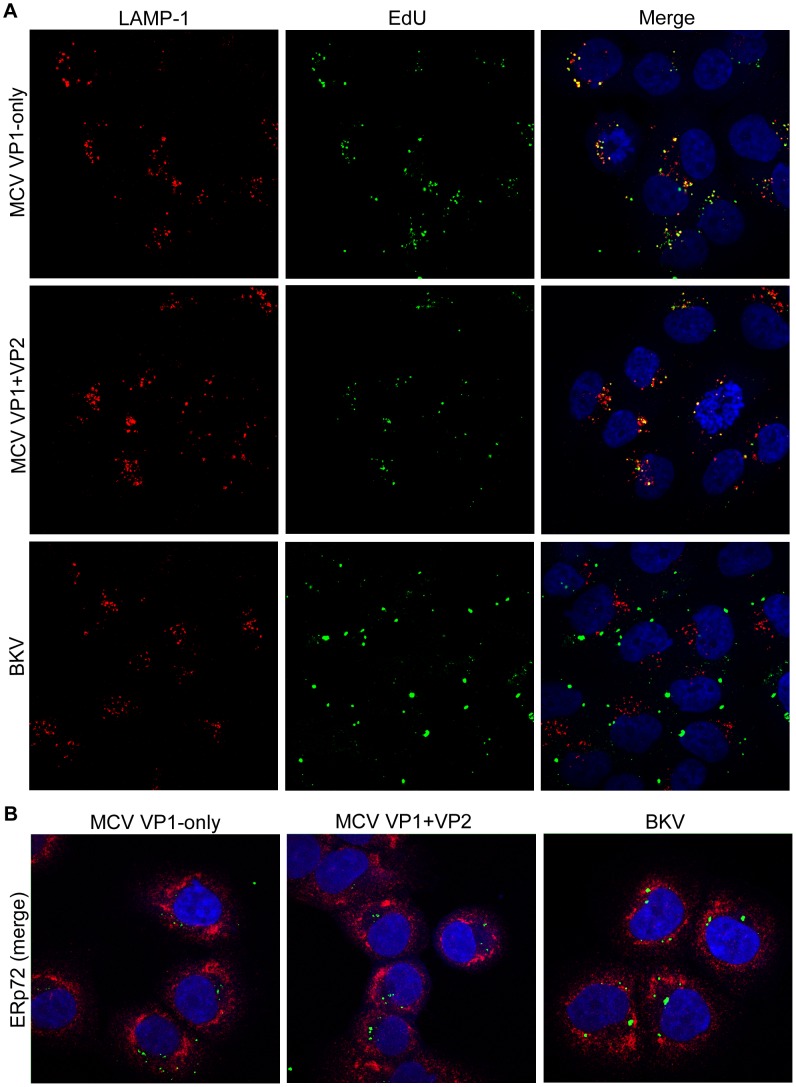
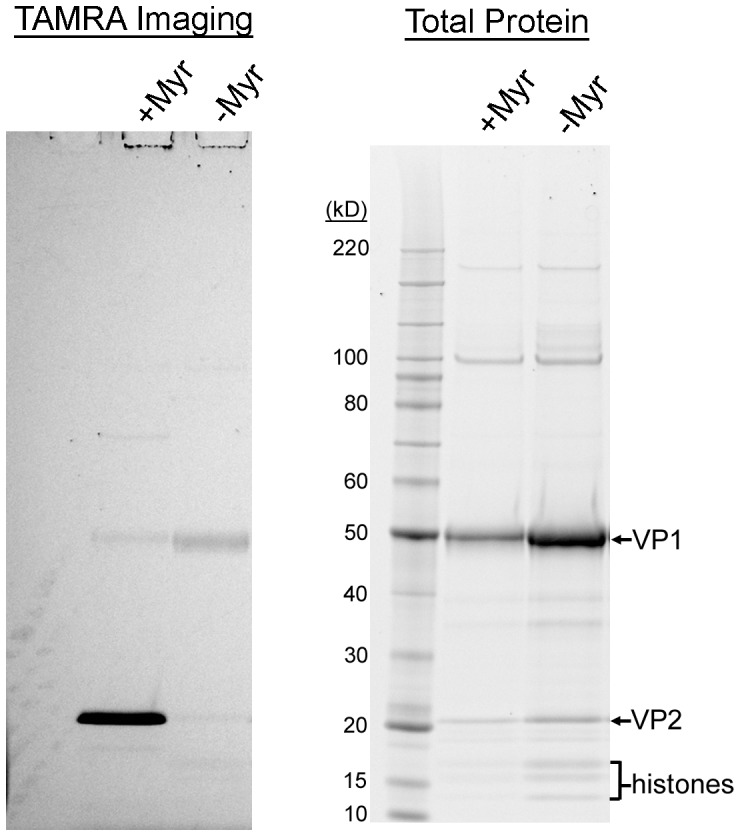
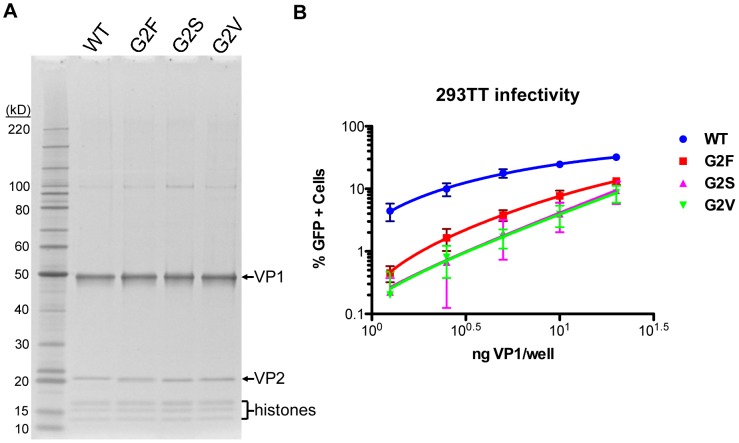
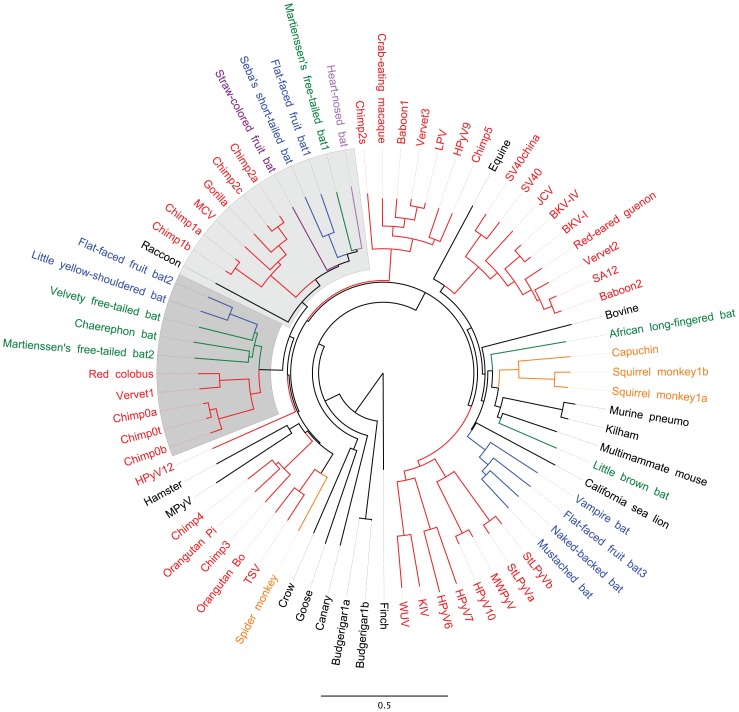
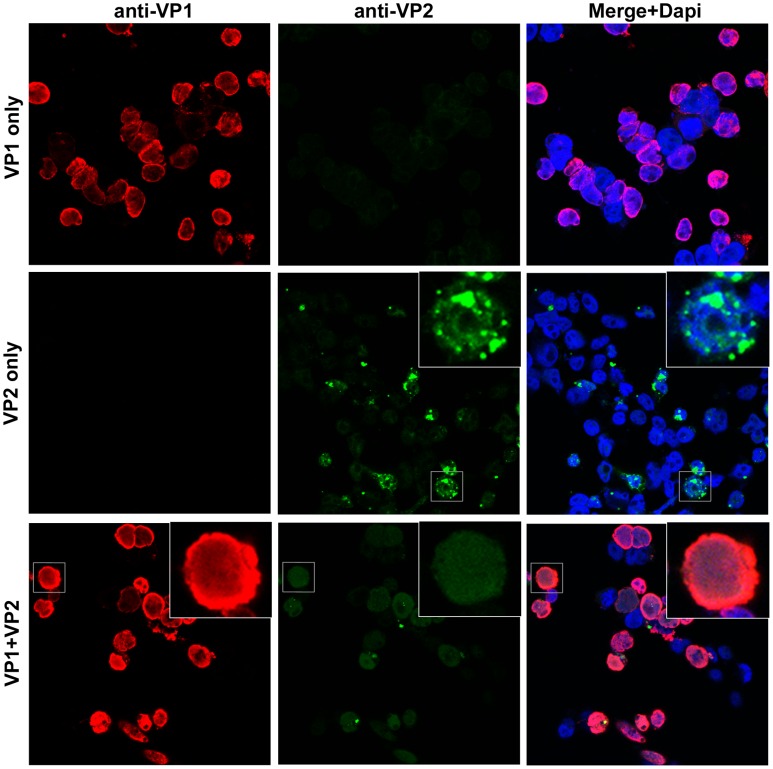
The surface of polyomavirus virions is composed of pentameric knobs of the major capsid protein, VP1. In previously studied polyomavirus species, such as SV40, two interior capsid proteins, VP2 and VP3, emerge from the virion to play important roles during the infectious entry process. Translation of the VP3 protein initiates at a highly conserved Met-Ala-Leu motif within the VP2 open reading frame. Phylogenetic analyses indicate that Merkel cell polyomavirus (MCV or MCPyV) is a member of a divergent clade of polyomaviruses that lack the conserved VP3 N-terminal motif. Consistent with this observation, we show that VP3 is not detectable in MCV-infected cells, VP3 is not found in native MCV virions, and mutation of possible alternative VP3-initiating methionine codons did not significantly affect MCV infectivity in culture. In contrast, VP2 knockout resulted in a >100-fold decrease in native MCV infectivity, despite normal virion assembly, viral DNA packaging, and cell attachment. Although pseudovirus-based experiments confirmed that VP2 plays an essential role for infection of some cell lines, other cell lines were readily transduced by pseudovirions lacking VP2. In cell lines where VP2 was needed for efficient infectious entry, the presence of a conserved myristoyl modification on the N-terminus of VP2 was important for its function. The results show that a single minor capsid protein, VP2, facilitates a post-attachment stage of MCV infectious entry into some, but not all, cell types.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
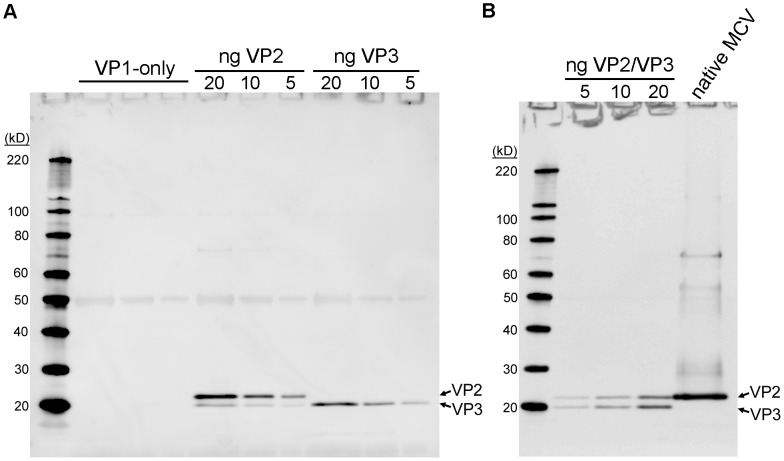
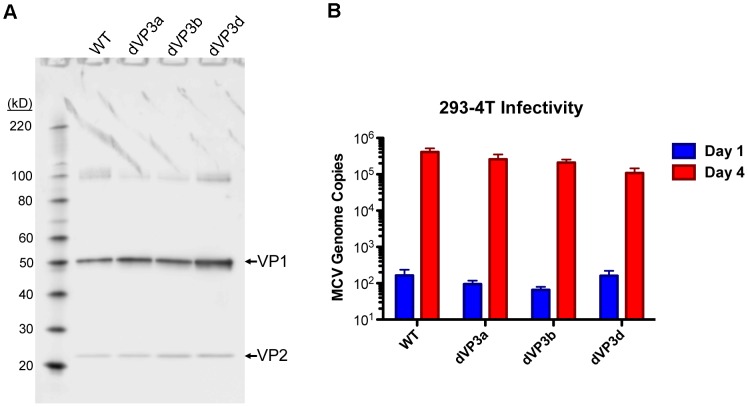
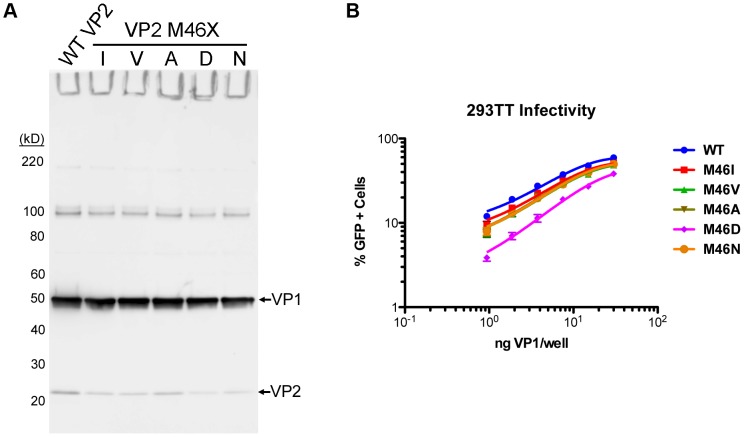
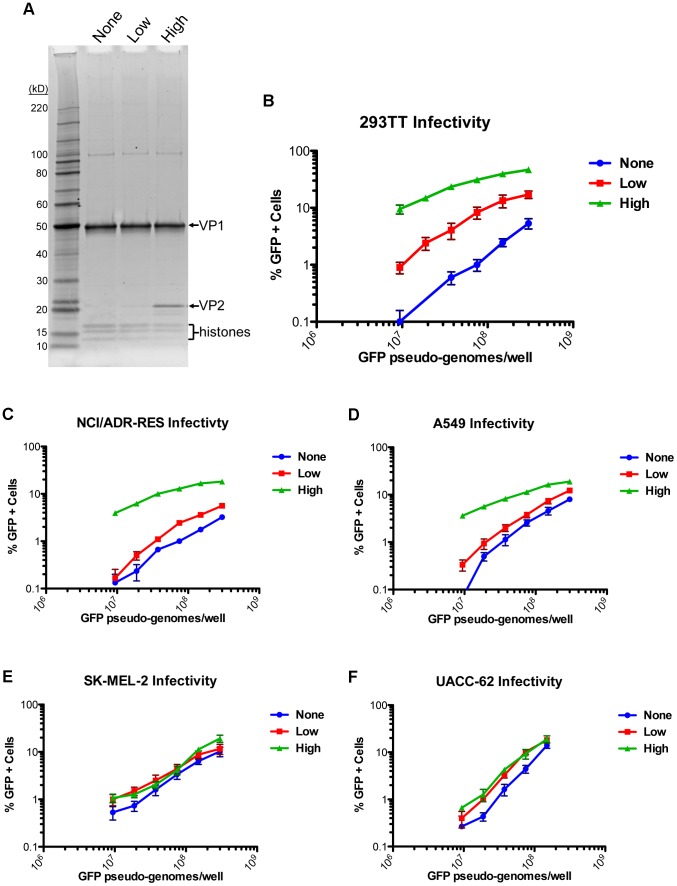
References
-
- Liddington RC, Yan Y, Moulai J, Sahli R, Benjamin TL, et al. (1991) Structure of simian virus 40 at 3.8-A resolution. Nature 354: 278–284. - PubMed
-
- Daniels R, Rusan NM, Wadsworth P, Hebert DN (2006) SV40 VP2 and VP3 insertion into ER membranes is controlled by the capsid protein VP1: implications for DNA translocation out of the ER. Mol Cell 24: 955–966. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
